

Diversity of innate immune recognition mechanism for bacterial polymeric meso-diaminopimelic acid-type peptidoglycan in insects
- PMID: 20702416
- PMCID: PMC2963372
- DOI: 10.1074/jbc.M110.144014
Diversity of innate immune recognition mechanism for bacterial polymeric meso-diaminopimelic acid-type peptidoglycan in insects
Abstract
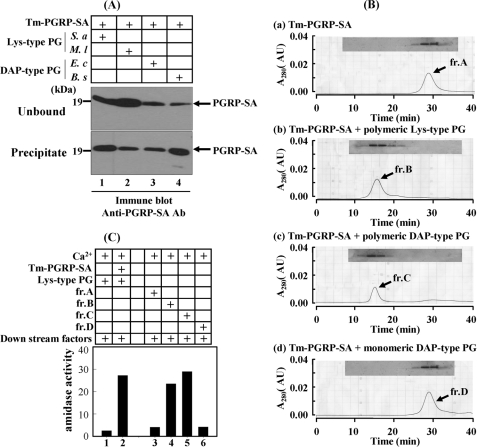
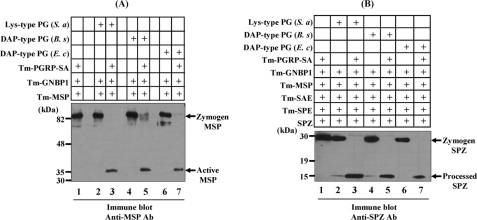
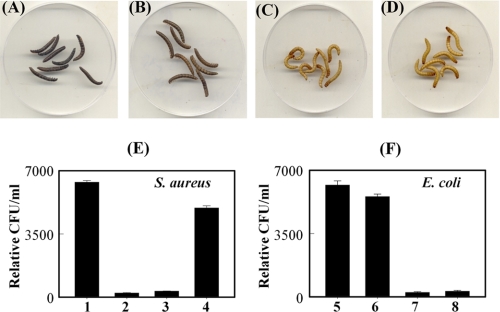
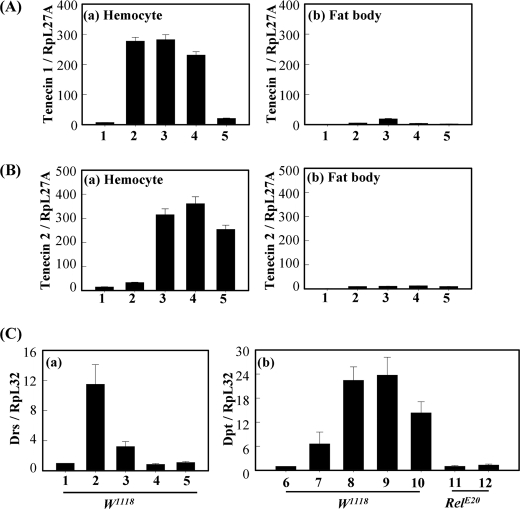
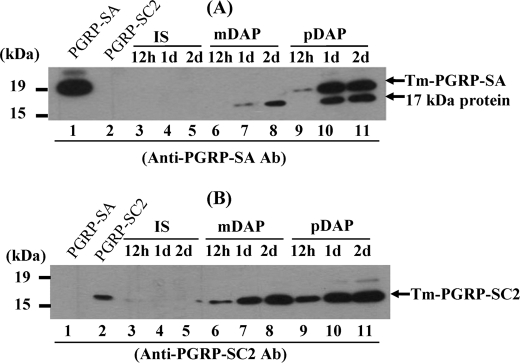
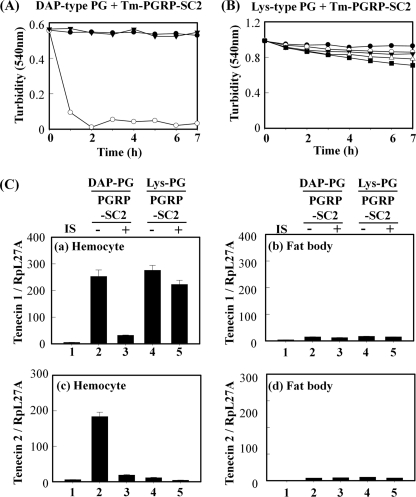
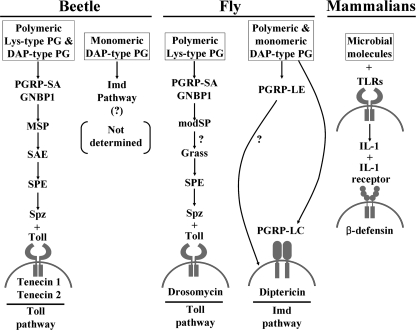
In Drosophila, the synthesis of antimicrobial peptides in response to microbial infections is under the control of the Toll and immune deficiency (Imd) signaling pathway. The Toll signaling pathway responds mainly to the lysine-type peptidoglycan of Gram-positive bacteria and fungal β-1,3-glucan, whereas the Imd pathway responds to the meso-diaminopimelic acid (DAP)-type peptidoglycan of Gram-negative bacteria and certain Gram-positive bacilli. Recently we determined the activation mechanism of a Toll signaling pathway biochemically using a large beetle, Tenebrio molitor. However, DAP-type peptidoglycan recognition mechanism and its signaling pathway are still unclear in the fly and beetle. Here, we show that polymeric DAP-type peptidoglycan, but not its monomeric form, formed a complex with Tenebrio peptidoglycan recognition protein-SA, and this complex activated the three-step proteolytic cascade to produce processed Spätzle, a Toll receptor ligand, and induced Drosophila defensin-like antimicrobial peptide in Tenebrio larvae similarly to polymeric lysine-type peptidoglycan. Monomeric DAP-type peptidoglycan induced Drosophila diptericin-like antimicrobial peptide in Tenebrio hemocytes. In addition, both polymeric and monomeric DAP-type peptidoglycans induced expression of Tenebrio peptidoglycan recognition protein-SC2, which is DAP-type peptidoglycan-selective N-acetylmuramyl-l-alanine amidase that functions as a DAP-type peptidoglycan scavenger, appearing to function as a negative regulator of the DAP-type peptidoglycan signaling by cleaving DAP-type peptidoglycan in Tenebrio larvae. Taken together, these results demonstrate that molecular recognition mechanism for polymeric DAP-type peptidoglycan is different between Tenebrio larvae and Drosophila adults, providing biochemical evidences of biological diversity of innate immune responses in insects.
Figures

Similar articles
-
Crystal structure of Drosophila PGRP-SD suggests binding to DAP-type but not lysine-type peptidoglycan.Mol Immunol. 2008 May;45(9):2521-30. doi: 10.1016/j.molimm.2008.01.015. Epub 2008 Mar 4. Mol Immunol. 2008. PMID: 18304640
-
Structural and Functional Analysis of PGRP-LC Indicates Exclusive Dap-Type PGN Binding in Bumblebees.Int J Mol Sci. 2020 Apr 1;21(7):2441. doi: 10.3390/ijms21072441. Int J Mol Sci. 2020. PMID: 32244587 Free PMC article.
-
A three-step proteolytic cascade mediates the activation of the peptidoglycan-induced toll pathway in an insect.J Biol Chem. 2008 Mar 21;283(12):7599-607. doi: 10.1074/jbc.M710216200. Epub 2008 Jan 14. J Biol Chem. 2008. PMID: 18195005
-
Autophagy in Tenebrio molitor Immunity: Conserved Antimicrobial Functions in Insect Defenses.Front Immunol. 2021 May 31;12:667664. doi: 10.3389/fimmu.2021.667664. eCollection 2021. Front Immunol. 2021. PMID: 34135896 Free PMC article. Review.
-
[Intra- and extracellular recognition of pathogens and activation of innate immunity].Yakugaku Zasshi. 2006 Dec;126(12):1213-8. doi: 10.1248/yakushi.126.1213. Yakugaku Zasshi. 2006. PMID: 17139146 Review. Japanese.
Cited by
-
Development on Citrus medica infected with 'Candidatus Liberibacter asiaticus' has sex-specific and -nonspecific impacts on adult Diaphorina citri and its endosymbionts.PLoS One. 2020 Oct 6;15(10):e0239771. doi: 10.1371/journal.pone.0239771. eCollection 2020. PLoS One. 2020. PMID: 33022020 Free PMC article.
-
Independent recognition of Staphylococcus aureus by two receptors for phagocytosis in Drosophila.J Biol Chem. 2012 Jun 22;287(26):21663-72. doi: 10.1074/jbc.M111.333807. Epub 2012 Apr 30. J Biol Chem. 2012. PMID: 22547074 Free PMC article.
-
TmSpz4 Plays an Important Role in Regulating the Production of Antimicrobial Peptides in Response to Escherichia coli and Candida albicans Infections.Int J Mol Sci. 2020 Mar 9;21(5):1878. doi: 10.3390/ijms21051878. Int J Mol Sci. 2020. PMID: 32182940 Free PMC article.
-
93-kDa twin-domain serine protease inhibitor (Serpin) has a regulatory function on the beetle Toll proteolytic signaling cascade.J Biol Chem. 2011 Oct 7;286(40):35087-95. doi: 10.1074/jbc.M111.277343. Epub 2011 Aug 23. J Biol Chem. 2011. PMID: 21862574 Free PMC article.
-
Prophenoloxidase activation and antimicrobial peptide expression induced by the recombinant microbe binding protein of Manduca sexta.Insect Biochem Mol Biol. 2017 Apr;83:35-43. doi: 10.1016/j.ibmb.2016.10.006. Epub 2016 Oct 29. Insect Biochem Mol Biol. 2017. PMID: 27989837 Free PMC article.
References
-
- Lemaitre B., Hoffmann J. (2007) Annu. Rev. Immunol. 25, 697–743 - PubMed
-
- Georgel P., Naitza S., Kappler C., Ferrandon D., Zachary D., Swimmer C., Kopczynski C., Duyk G., Reichhart J. M., Hoffmann J. A. (2001) Dev. Cell 1, 503–514 - PubMed
-
- Lemaitre B., Nicolas E., Michaut L., Reichhart J. M., Hoffmann J. A. (1996) Cell 86, 973–983 - PubMed
-
- Hedengren M., Asling B., Dushay M. S., Ando I., Ekengren S., Wihlborg M., Hultmark D. (1999) Mol. Cell 4, 827–837 - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
