

Conditions for mutation-order speciation
- PMID: 20702458
- PMCID: PMC3013408
- DOI: 10.1098/rspb.2010.1215
Conditions for mutation-order speciation
Abstract
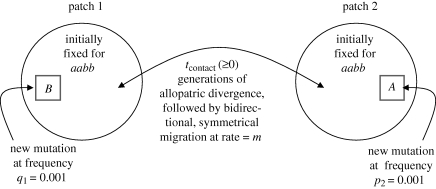
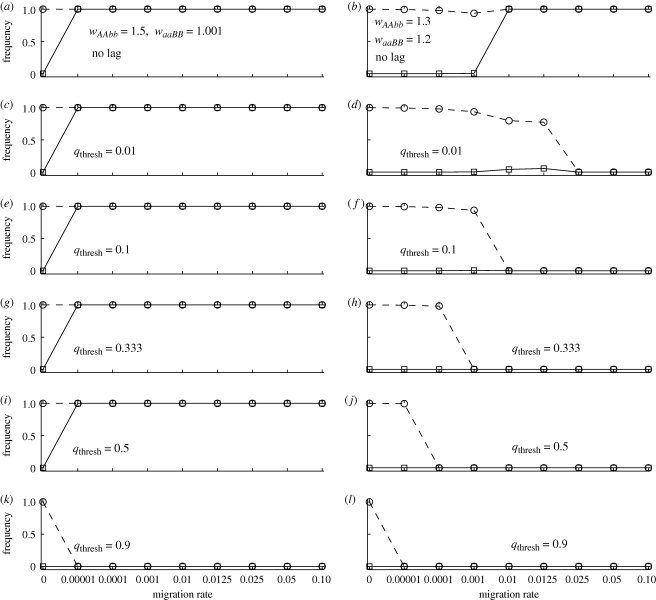
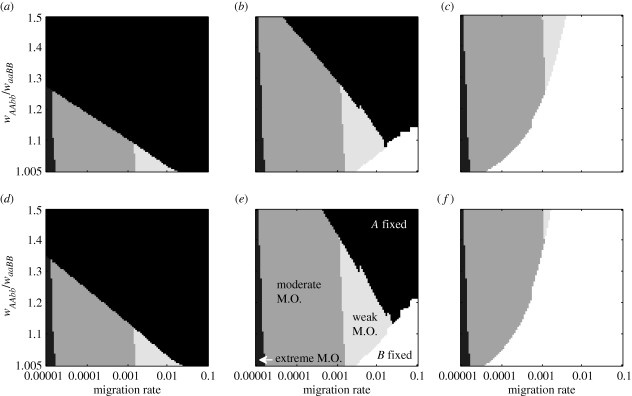
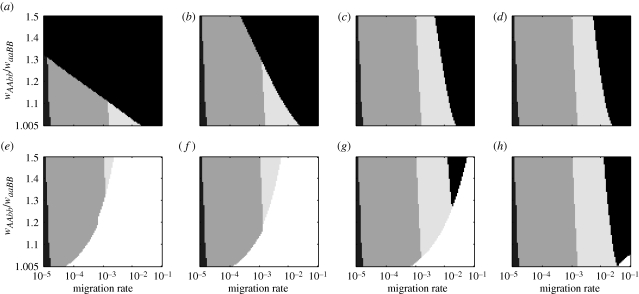
Two models for speciation via selection have been proposed. In the well-known model of 'ecological speciation', divergent natural selection between environments drives the evolution of reproductive isolation. In a second 'mutation-order' model, different, incompatible mutations (alleles) fix in different populations adapting to the same selective pressure. How to demonstrate mutation-order speciation has been unclear, although it has been argued that it can be ruled out when gene flow occurs because the same, most advantageous allele will fix in all populations. However, quantitative examination of the interaction of factors influencing the likelihood of mutation-order speciation is lacking. We used simulation models to study how gene flow, hybrid incompatibility, selective advantage, timing of origination of new mutations and an initial period of allopatric differentiation affect population divergence via the mutation-order process. We find that at least some population divergence can occur under a reasonably wide range of conditions, even with moderate gene flow. However, strong divergence (e.g. fixation of different alleles in different populations) requires very low gene flow, and is promoted when (i) incompatible mutations have similar fitness advantages, (ii) less fit mutations arise slightly earlier in evolutionary time than more fit alternatives, and (iii) allopatric divergence occurs prior to secondary contact.
Figures
Similar articles
-
Mutation-order divergence by sexual selection: diversification of sexual signals in similar environments as a first step in speciation.Ecol Lett. 2014 Sep;17(9):1053-66. doi: 10.1111/ele.12313. Epub 2014 Jun 18. Ecol Lett. 2014. PMID: 24943881
-
Insights from Fisher's geometric model on the likelihood of speciation under different histories of environmental change.Evolution. 2020 Aug;74(8):1603-1619. doi: 10.1111/evo.14032. Epub 2020 Jun 25. Evolution. 2020. PMID: 32542705
-
Evidence for ecological speciation and its alternative.Science. 2009 Feb 6;323(5915):737-41. doi: 10.1126/science.1160006. Science. 2009. PMID: 19197053 Review.
-
The existence of species rests on a metastable equilibrium between inbreeding and outbreeding. An essay on the close relationship between speciation, inbreeding and recessive mutations.Biol Direct. 2011 Dec 9;6:62. doi: 10.1186/1745-6150-6-62. Biol Direct. 2011. PMID: 22152499 Free PMC article.
-
Genetics and ecological speciation.Proc Natl Acad Sci U S A. 2009 Jun 16;106 Suppl 1(Suppl 1):9955-62. doi: 10.1073/pnas.0901264106. Epub 2009 Jun 15. Proc Natl Acad Sci U S A. 2009. PMID: 19528639 Free PMC article. Review.
Cited by
-
Unifying models of dialect spread and extinction using surface tension dynamics.R Soc Open Sci. 2018 Jan 3;5(1):171446. doi: 10.1098/rsos.171446. eCollection 2018 Jan. R Soc Open Sci. 2018. PMID: 29410847 Free PMC article.
-
Reproductive isolation between phylogeographic lineages scales with divergence.Proc Biol Sci. 2013 Oct 9;280(1772):20132246. doi: 10.1098/rspb.2013.2246. Print 2013 Dec 7. Proc Biol Sci. 2013. PMID: 24107536 Free PMC article.
-
Geographic and Ecological Dimensions of Host Plant-Associated Genetic Differentiation and Speciation in the Rhagoletis cingulata (Diptera: Tephritidae) Sibling Species Group.Insects. 2019 Aug 29;10(9):275. doi: 10.3390/insects10090275. Insects. 2019. PMID: 31470668 Free PMC article.
-
Initiation of speciation across multiple dimensions in a rock-restricted, tropical lizard.Mol Ecol. 2023 Feb;32(3):680-695. doi: 10.1111/mec.16787. Epub 2022 Nov 27. Mol Ecol. 2023. PMID: 36394360 Free PMC article.
-
Coupling of twelve putative chromosomal inversions maintains a strong barrier to gene flow between snail ecotypes.Evol Lett. 2024 Apr 23;8(4):575-586. doi: 10.1093/evlett/qrae014. eCollection 2024 Aug. Evol Lett. 2024. PMID: 39479507 Free PMC article.
References
-
- Coyne J. A., Orr H. A. 2004. Speciation. Sunderland, MA: Sinauer Associates, Inc
-
- Gavrilets S. 2004. Fitness landscapes and the origin of species. Princeton, NJ: Princeton University Press
-
- Funk D. J., Nosil P., Etges B. 2006. Ecological divergence exhibits consistently positive associations with reproductive isolation across disparate taxa. Proc. Natl Acad. Sci. USA 103, 3209–321310.1073/pnas.0508653103 (doi:10.1073/pnas.0508653103) - DOI - DOI - PMC - PubMed
-
- Schluter D. 2009. Evidence for ecological speciation and its alternative. Science 323, 737–74110.1126/science.1160006 (doi:10.1126/science.1160006) - DOI - DOI - PubMed
-
- Sobel J. M., Chen G. F., Watt L. R., Schemske D. W. 2010. The biology of speciation. Evolution 64, 295–31510.1111/j.1558-5646.2009.00877.x (doi:10.1111/j.1558-5646.2009.00877.x) - DOI - DOI - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
