

A congruent solution to arthropod phylogeny: phylogenomics, microRNAs and morphology support monophyletic Mandibulata
- PMID: 20702459
- PMCID: PMC3013382
- DOI: 10.1098/rspb.2010.0590
A congruent solution to arthropod phylogeny: phylogenomics, microRNAs and morphology support monophyletic Mandibulata
Abstract
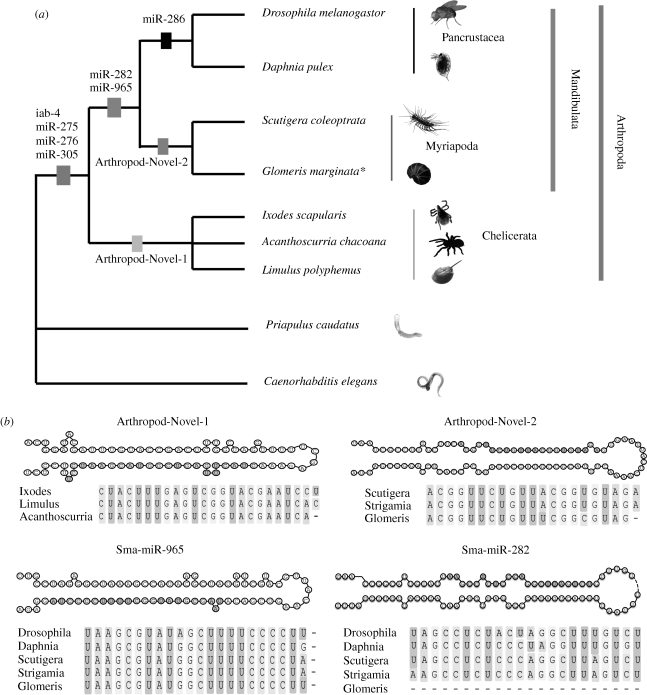
While a unique origin of the euarthropods is well established, relationships between the four euarthropod classes-chelicerates, myriapods, crustaceans and hexapods-are less clear. Unsolved questions include the position of myriapods, the monophyletic origin of chelicerates, and the validity of the close relationship of euarthropods to tardigrades and onychophorans. Morphology predicts that myriapods, insects and crustaceans form a monophyletic group, the Mandibulata, which has been contradicted by many molecular studies that support an alternative Myriochelata hypothesis (Myriapoda plus Chelicerata). Because of the conflicting insights from published molecular datasets, evidence from nuclear-coding genes needs corroboration from independent data to define the relationships among major nodes in the euarthropod tree. Here, we address this issue by analysing two independent molecular datasets: a phylogenomic dataset of 198 protein-coding genes including new sequences for myriapods, and novel microRNA complements sampled from all major arthropod lineages. Our phylogenomic analyses strongly support Mandibulata, and show that Myriochelata is a tree-reconstruction artefact caused by saturation and long-branch attraction. The analysis of the microRNA dataset corroborates the Mandibulata, showing that the microRNAs miR-965 and miR-282 are present and expressed in all mandibulate species sampled, but not in the chelicerates. Mandibulata is further supported by the phylogenetic analysis of a comprehensive morphological dataset covering living and fossil arthropods, and including recently proposed, putative apomorphies of Myriochelata. Our phylogenomic analyses also provide strong support for the inclusion of pycnogonids in a monophyletic Chelicerata, a paraphyletic Cycloneuralia, and a common origin of Arthropoda (tardigrades, onychophorans and arthropods), suggesting that previous phylogenies grouping tardigrades and nematodes may also have been subject to tree-reconstruction artefacts.
Figures




References
-
- Telford M. J., Bourlat S. J., Economou A. D., Papillon D., Rota-Strabelli O. 2008. The evolution of the Ecdysozoa. Phil. Trans. R. Soc. B 363, 1529–153710.1098/rstb.2007.2243 (doi:10.1098/rstb.2007.2243) - DOI - DOI - PMC - PubMed
-
- Edgecombe G. D. 2010. Arthropod phylogeny: an overview from the perspectives of morphology, molecular data and the fossil record. Arthropod Struct. Dev. 39, 74–8710.1016/j.asd.2009.10.002 (doi:10.1016/j.asd.2009.10.002) - DOI - DOI - PubMed
-
- Budd G. E., Telford M. J. 2009. The origin and evolution of arthropods. Nature 417, 812–81710.1038/nature07890 (doi:10.1038/nature07890) - DOI - DOI - PubMed
-
- Nielsen C. 1995. Animal evolution. Interrelationships of the living phyla, 2nd edn. Oxford, UK: Oxford University Press
-
- Telford M. J., Thomas R. H. 1995. Demise of the Atelocerata? Nature 376, 123–12410.1038/376123a0 (doi:10.1038/376123a0) - DOI - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
