

An SNP in an ultraconserved regulatory element affects Dlx5/Dlx6 regulation in the forebrain
- PMID: 20702565
- PMCID: PMC3188590
- DOI: 10.1242/dev.051052
An SNP in an ultraconserved regulatory element affects Dlx5/Dlx6 regulation in the forebrain
Abstract
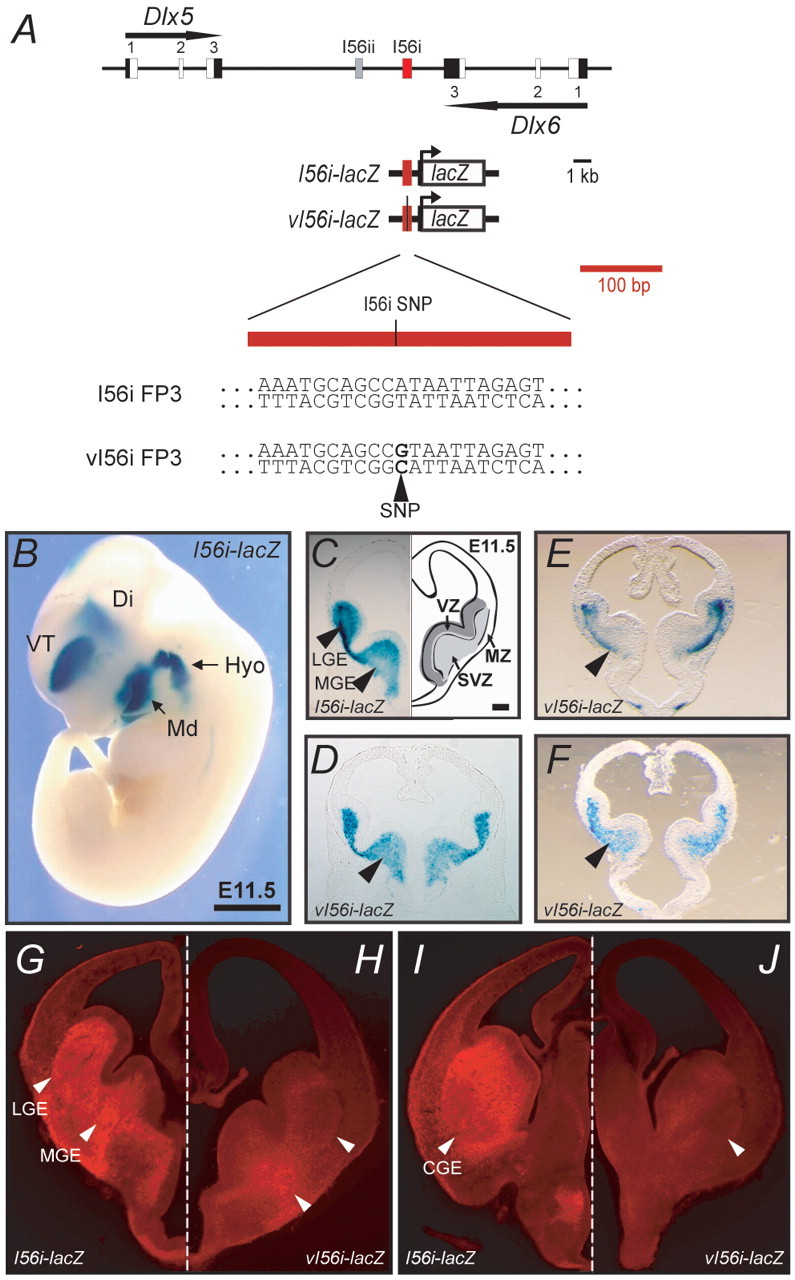
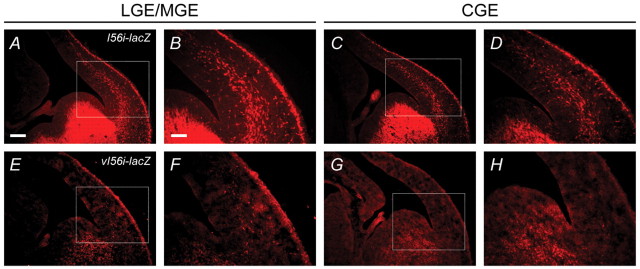
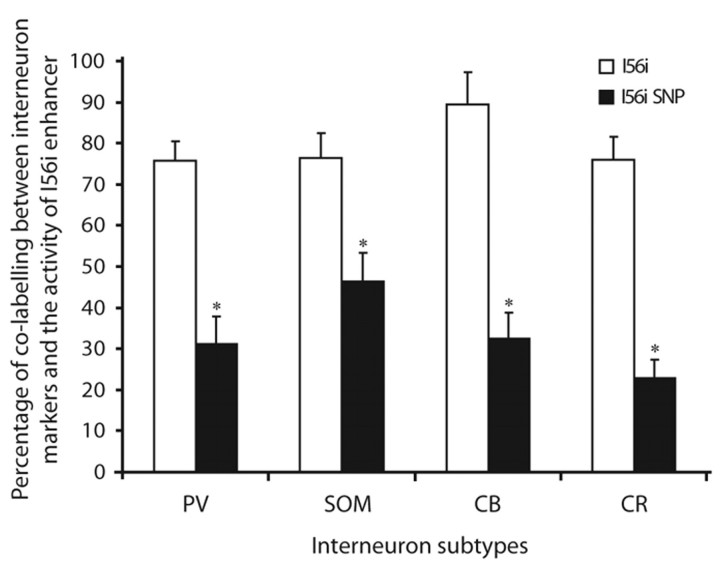
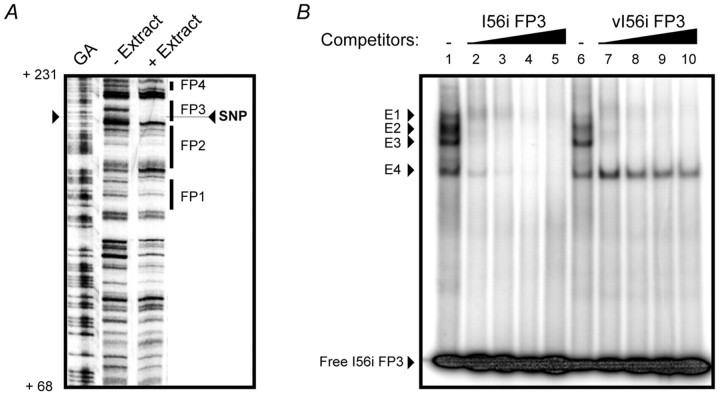
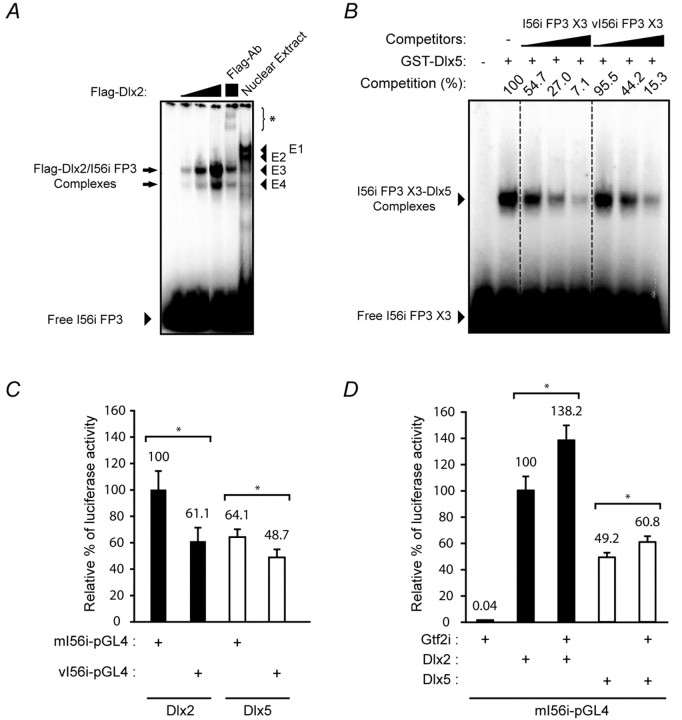
Dlx homeobox genes play a crucial role in the migration and differentiation of the subpallial precursor cells that give rise to various subtypes of gamma-aminobutyric acid (GABA)-expressing neurons of the forebrain, including local-circuit cortical interneurons. Aberrant development of GABAergic interneurons has been linked to several neurodevelopmental disorders, including epilepsy, schizophrenia, Rett syndrome and autism. Here, we report in mice that a single-nucleotide polymorphism (SNP) found in an autistic proband falls within a functional protein binding site in an ultraconserved cis-regulatory element. This element, I56i, is involved in regulating Dlx5/Dlx6 homeobox gene expression in the developing forebrain. We show that the SNP results in reduced I56i activity, predominantly in the medial and caudal ganglionic eminences and in streams of neurons tangentially migrating to the cortex. Reduced activity is also observed in GABAergic interneurons of the adult somatosensory cortex. The SNP affects the affinity of Dlx proteins for their binding site in vitro and reduces the transcriptional activation of the enhancer by Dlx proteins. Affinity purification using I56i sequences led to the identification of a novel regulator of Dlx gene expression, general transcription factor 2 I (Gtf2i), which is among the genes most often deleted in Williams-Beuren syndrome, a neurodevelopmental disorder. This study illustrates the clear functional consequences of a single nucleotide variation in an ultraconserved non-coding sequence in the context of developmental abnormalities associated with disease.
Figures

Similar articles
-
A highly conserved enhancer in the Dlx5/Dlx6 intergenic region is the site of cross-regulatory interactions between Dlx genes in the embryonic forebrain.J Neurosci. 2000 Jan 15;20(2):709-21. doi: 10.1523/JNEUROSCI.20-02-00709.2000. J Neurosci. 2000. PMID: 10632600 Free PMC article.
-
Distinct cis-regulatory elements from the Dlx1/Dlx2 locus mark different progenitor cell populations in the ganglionic eminences and different subtypes of adult cortical interneurons.J Neurosci. 2007 May 9;27(19):5012-22. doi: 10.1523/JNEUROSCI.4725-06.2007. J Neurosci. 2007. PMID: 17494687 Free PMC article.
-
Activity of dlx5a/dlx6a regulatory elements during zebrafish GABAergic neuron development.Int J Dev Neurosci. 2011 Nov;29(7):681-91. doi: 10.1016/j.ijdevneu.2011.06.005. Epub 2011 Jun 23. Int J Dev Neurosci. 2011. PMID: 21723936
-
DLX genes and proteins in mammalian forebrain development.Development. 2024 Jun 1;151(11):dev202684. doi: 10.1242/dev.202684. Epub 2024 May 31. Development. 2024. PMID: 38819455 Free PMC article. Review.
-
Homeobox genes in vertebrate forebrain development and disease.Clin Genet. 2008 Mar;73(3):212-26. doi: 10.1111/j.1399-0004.2008.00967.x. Epub 2008 Jan 31. Clin Genet. 2008. PMID: 18241223 Review.
Cited by
-
Dlx5/6 Expression Levels in Mouse GABAergic Neurons Regulate Adult Parvalbumin Neuronal Density and Anxiety/Compulsive Behaviours.Cells. 2022 May 25;11(11):1739. doi: 10.3390/cells11111739. Cells. 2022. PMID: 35681437 Free PMC article.
-
Ultraconserved Elements Occupy Specific Arenas of Three-Dimensional Mammalian Genome Organization.Cell Rep. 2018 Jul 10;24(2):479-488. doi: 10.1016/j.celrep.2018.06.031. Cell Rep. 2018. PMID: 29996107 Free PMC article.
-
DLX5/6 GABAergic Expression Affects Social Vocalization: Implications for Human Evolution.Mol Biol Evol. 2021 Oct 27;38(11):4748-4764. doi: 10.1093/molbev/msab181. Mol Biol Evol. 2021. PMID: 34132815 Free PMC article.
-
GC-biased gene conversion drives accelerated evolution of ultraconserved elements in mammalian and avian genomes.Genome Res. 2023 Oct;33(10):1673-1689. doi: 10.1101/gr.277784.123. Epub 2023 Oct 26. Genome Res. 2023. PMID: 37884342 Free PMC article.
-
Dlx1 and Dlx2 Promote Interneuron GABA Synthesis, Synaptogenesis, and Dendritogenesis.Cereb Cortex. 2018 Nov 1;28(11):3797-3815. doi: 10.1093/cercor/bhx241. Cereb Cortex. 2018. PMID: 29028947 Free PMC article.
References
-
- Anderson S. A., Eisenstat D. D., Shi L., Rubenstein J. L. R. (1997a). Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science 278, 474-476 - PubMed
-
- Anderson S. A., Qiu M., Bulfone A., Eisenstat D. D., Meneses J., Pedersen R., Rubenstein J. L. R. (1997b). Mutations of the homeobox genes Dlx-1 and Dlx-2 disrupt the striatal subventricular zone and differentiation of late-born striatal neurons. Neuron 19, 27-37 - PubMed
-
- Bejerano G., Pheasant M., Makunin I., Stephen S., Kent W. J., Mattick J. S., Haussler D. (2004). Ultraconserved elements in the human genome. Science 304, 1321-1325 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
