

"Depupylation" of prokaryotic ubiquitin-like protein from mycobacterial proteasome substrates
- PMID: 20705495
- PMCID: PMC2939144
- DOI: 10.1016/j.molcel.2010.07.019
"Depupylation" of prokaryotic ubiquitin-like protein from mycobacterial proteasome substrates
Abstract
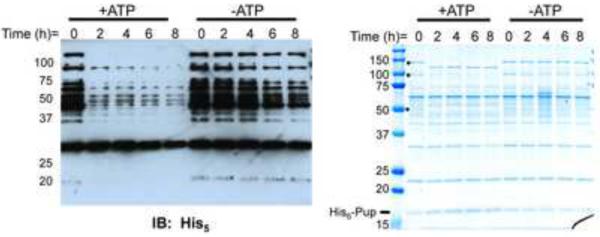
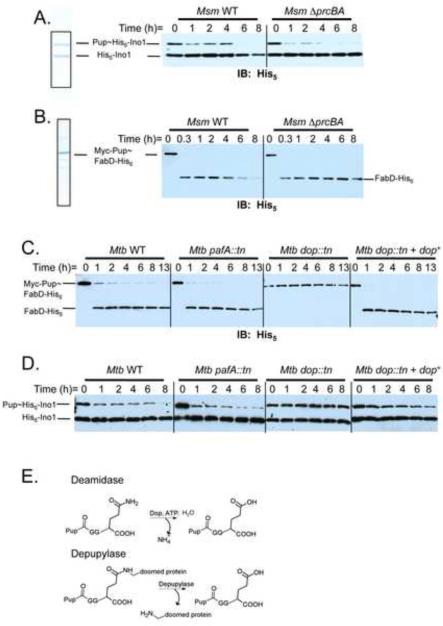
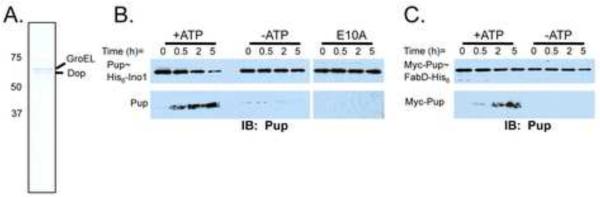
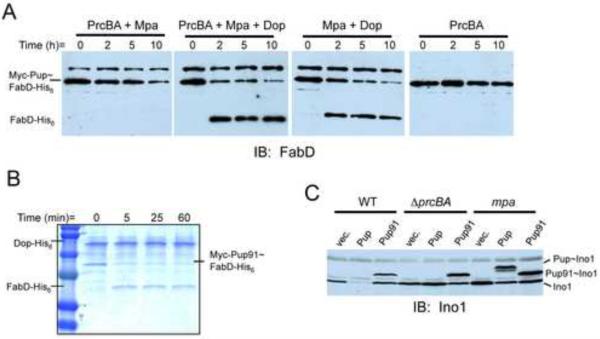
Ubiquitin (Ub) provides the recognition and specificity required to deliver proteins to the eukaryotic proteasome for destruction. Prokaryotic ubiquitin-like protein (Pup) is functionally analogous to Ub in Mycobacterium tuberculosis (Mtb), as it dooms proteins to the Mtb proteasome. Studies suggest that Pup and Ub do not share similar mechanisms of activation and conjugation to target proteins. Dop (deamidase of Pup; Mtb Rv2112c/MT2172) deamidates the C-terminal glutamine of Pup to glutamate, preparing it for ligation to target proteins by proteasome accessory factor A (PafA). While studies have shed light on the conjugation of Pup to proteins, it was not known if Pup could be removed from substrates in a manner analogous to the deconjugation of Ub from eukaryotic proteins. Here, we show that Mycobacteria have a "depupylase" activity provided by Dop. The discovery of a depupylase strengthens the parallels between the Pup- and Ub-tagging systems of prokaryotes and eukaryotes, respectively.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Brunger AT, DeLaBarre B. NSF and p97/VCP: similar at first, different at last. FEBS Lett. 2003;555:126–133. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
