

A screening pipeline for antiparasitic agents targeting cryptosporidium inosine monophosphate dehydrogenase
- PMID: 20706578
- PMCID: PMC2919388
- DOI: 10.1371/journal.pntd.0000794
A screening pipeline for antiparasitic agents targeting cryptosporidium inosine monophosphate dehydrogenase
Abstract
Background: The protozoan parasite Cryptosporidium parvum is responsible for significant disease burden among children in developing countries. In addition Cryptosporidiosis can result in chronic and life-threatening enteritis in AIDS patients, and the currently available drugs lack efficacy in treating these severe conditions. The discovery and development of novel anti-cryptosporidial therapeutics has been hampered by the poor experimental tractability of this pathogen. While the genome sequencing effort has identified several intriguing new targets including a unique inosine monophosphate dehydrogenase (IMPDH), pursuing these targets and testing inhibitors has been frustratingly difficult.
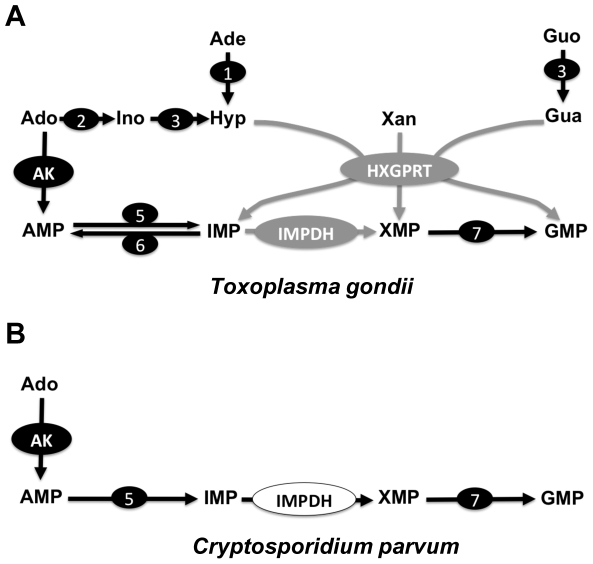
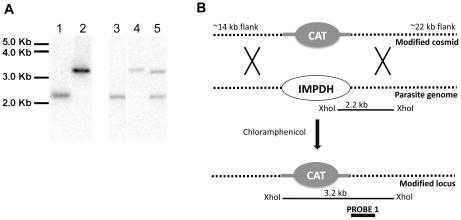
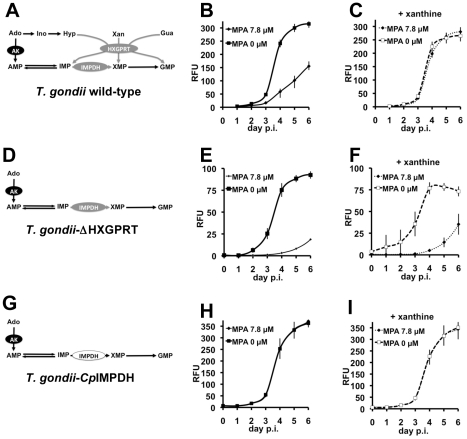
Methodology and principal findings: Here we have developed a pipeline of tools to accelerate the in vivo screening of inhibitors of C. parvum IMPDH. We have genetically engineered the related parasite Toxoplasma gondii to serve as a model of C. parvum infection as the first screen. This assay provides crucial target validation and a large signal window that is currently not possible in assays involving C. parvum. To further develop compounds that pass this first filter, we established a fluorescence-based assay of host cell proliferation, and a C. parvum growth assay that utilizes automated high-content imaging analysis for enhanced throughput.
Conclusions and significance: We have used these assays to evaluate C. parvum IMPDH inhibitors emerging from our ongoing medicinal chemistry effort and have identified a subset of 1,2,3-triazole ethers that exhibit excellent in vivo selectivity in the T. gondii model and improved anti-cryptosporidial activity.
Conflict of interest statement
Some of the authors of this paper are also listed on a patent application on the compounds described in this paper. The authors note that their primary goal in protecting the IP for the compounds is to ensure that they retain a chance to become actual drugs in the future. Unresolved IP often prevents further development of drugs. Nonetheless, the patent constitutes a potential competing interest.
Figures





Similar articles
-
Triazole inhibitors of Cryptosporidium parvum inosine 5'-monophosphate dehydrogenase.J Med Chem. 2009 Aug 13;52(15):4623-30. doi: 10.1021/jm900410u. J Med Chem. 2009. PMID: 19624136 Free PMC article.
-
Phthalazinone inhibitors of inosine-5'-monophosphate dehydrogenase from Cryptosporidium parvum.Bioorg Med Chem Lett. 2013 Feb 15;23(4):1004-7. doi: 10.1016/j.bmcl.2012.12.037. Epub 2012 Dec 27. Bioorg Med Chem Lett. 2013. PMID: 23324406 Free PMC article.
-
Effect of urea and squaramide IMPDH inhibitors on C. parvum: in vitro trial design impacts the assessment of drug efficacy.Int J Parasitol Drugs Drug Resist. 2025 Aug;28:100592. doi: 10.1016/j.ijpddr.2025.100592. Epub 2025 Apr 15. Int J Parasitol Drugs Drug Resist. 2025. PMID: 40319744 Free PMC article.
-
Reprofiled drug targets ancient protozoans: drug discovery for parasitic diarrheal diseases.Gut Microbes. 2013 Jan-Feb;4(1):66-71. doi: 10.4161/gmic.22596. Epub 2012 Nov 8. Gut Microbes. 2013. PMID: 23137963 Free PMC article. Review.
-
Inosine 5'-Monophosphate Dehydrogenase (IMPDH) as a Potential Target for the Development of a New Generation of Antiprotozoan Agents.Mini Rev Med Chem. 2018;18(8):656-671. doi: 10.2174/1389557516666160620065558. Mini Rev Med Chem. 2018. PMID: 27334467 Review.
Cited by
-
Optimization of benzoxazole-based inhibitors of Cryptosporidium parvum inosine 5'-monophosphate dehydrogenase.J Med Chem. 2013 May 23;56(10):4028-43. doi: 10.1021/jm400241j. Epub 2013 May 13. J Med Chem. 2013. PMID: 23668331 Free PMC article.
-
Synthesis and biological evaluation of ursolic acid derivatives bearing triazole moieties as potential anti-Toxoplasma gondii agents.J Enzyme Inhib Med Chem. 2019 Dec;34(1):761-772. doi: 10.1080/14756366.2019.1584622. J Enzyme Inhib Med Chem. 2019. PMID: 30836795 Free PMC article.
-
Mycobacterium tuberculosis IMPDH in Complexes with Substrates, Products and Antitubercular Compounds.PLoS One. 2015 Oct 6;10(10):e0138976. doi: 10.1371/journal.pone.0138976. eCollection 2015. PLoS One. 2015. PMID: 26440283 Free PMC article.
-
Mycophenolic anilides as broad specificity inosine-5'-monophosphate dehydrogenase (IMPDH) inhibitors.Bioorg Med Chem Lett. 2020 Dec 15;30(24):127543. doi: 10.1016/j.bmcl.2020.127543. Epub 2020 Sep 12. Bioorg Med Chem Lett. 2020. PMID: 32931912 Free PMC article.
-
Genetic modification of the diarrhoeal pathogen Cryptosporidium parvum.Nature. 2015 Jul 23;523(7561):477-80. doi: 10.1038/nature14651. Epub 2015 Jul 15. Nature. 2015. PMID: 26176919 Free PMC article.
References
-
- Snelling WJ, Xiao L, Ortega-Pierres G, Lowery CJ, Moore JE, et al. Cryptosporidiosis in developing countries. J Infect Dev Ctries. 2007;1:242–256. - PubMed
-
- Khan WA, Rogers KA, Karim MM, Ahmed S, Hibberd PL, et al. Cryptosporidiosis among Bangladeshi children with diarrhea: a prospective, matched, case-control study of clinical features, epidemiology and systemic antibody responses. Am J Trop Med Hyg. 2004;71:412–419. - PubMed
-
- Lima AA, Fang G, Schorling JB, de Albuquerque L, McAuliffe JF, et al. Persistent diarrhea in northeast Brazil: etiologies and interactions with malnutrition. Acta Paediatr Suppl. 1992;381:39–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
