

Hemocyte-secreted type IV collagen enhances BMP signaling to guide renal tubule morphogenesis in Drosophila
- PMID: 20708591
- PMCID: PMC2941037
- DOI: 10.1016/j.devcel.2010.07.019
Hemocyte-secreted type IV collagen enhances BMP signaling to guide renal tubule morphogenesis in Drosophila
Abstract
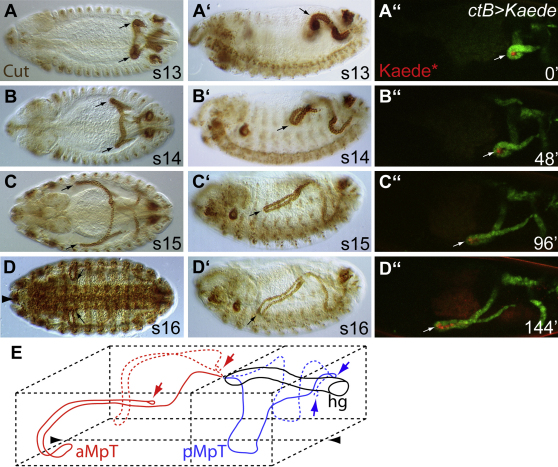
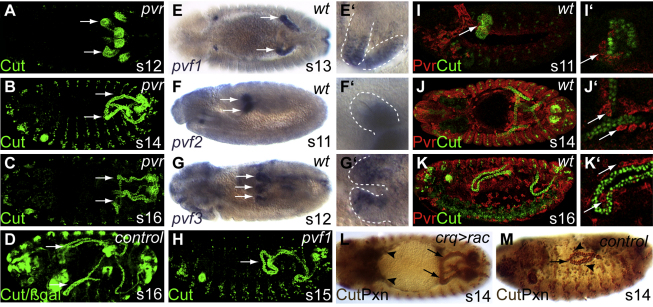
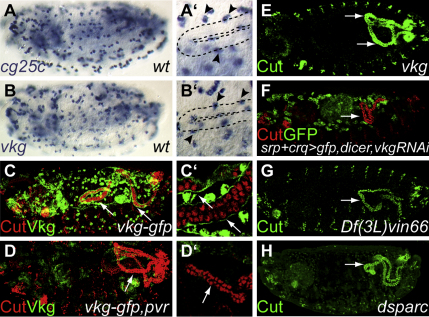
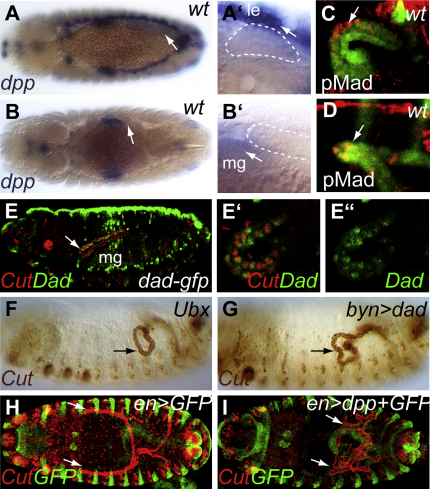
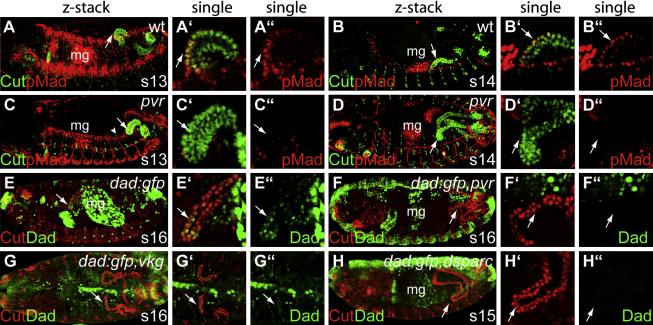
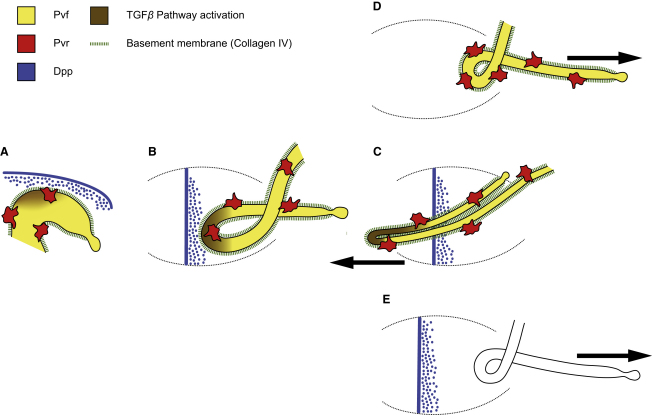
Details of the mechanisms that determine the shape and positioning of organs in the body cavity remain largely obscure. We show that stereotypic positioning of outgrowing Drosophila renal tubules depends on signaling in a subset of tubule cells and results from enhanced sensitivity to guidance signals by targeted matrix deposition. VEGF/PDGF ligands from the tubules attract hemocytes, which secrete components of the basement membrane to ensheath them. Collagen IV sensitizes tubule cells to localized BMP guidance cues. Signaling results in pathway activation in a subset of tubule cells that lead outgrowth through the body cavity. Failure of hemocyte migration, loss of collagen IV, or abrogation of BMP signaling results in tubule misrouting and defective organ shape and positioning. Such regulated interplay between cell-cell and cell-matrix interactions is likely to have wide relevance in organogenesis and congenital disease.
2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Affolter M., Basler K. The Decapentaplegic morphogen gradient: from pattern formation to growth regulation. Nat. Rev. Genet. 2007;8:663–674. - PubMed
-
- Affolter M., Nellen D., Nussbaumer U., Basler K. Multiple requirements for the receptor serine/threonine kinase thick veins reveal novel functions of TGF beta homologs during Drosophila embryogenesis. Development. 1994;120:3105–3117. - PubMed
-
- Arora K., Dai H., Kazuko S.G., Jamal J., O'Connor M.B., Letsou A., Warrior R. The Drosophila schnurri gene acts in the Dpp/TGF beta signaling pathway and encodes a transcription factor homologous to the human MBP family. Cell. 1995;81:781–790. - PubMed
-
- Baena-Lopez L.A., Baonza A., Garcia-Bellido A. The orientation of cell divisions determines the shape of Drosophila organs. Curr. Biol. 2005;15:1640–1644. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
