

The cystic fibrosis transmembrane conductance regulator impedes proteolytic stimulation of the epithelial Na+ channel
- PMID: 20709758
- PMCID: PMC2952223
- DOI: 10.1074/jbc.M110.155259
The cystic fibrosis transmembrane conductance regulator impedes proteolytic stimulation of the epithelial Na+ channel
Abstract
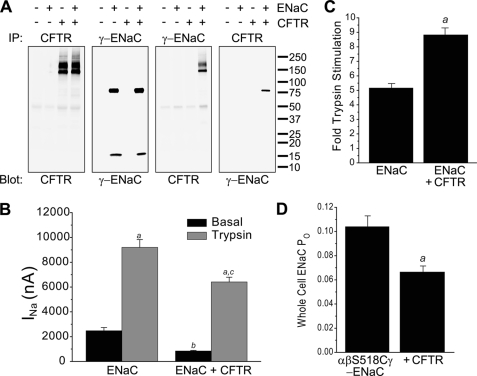
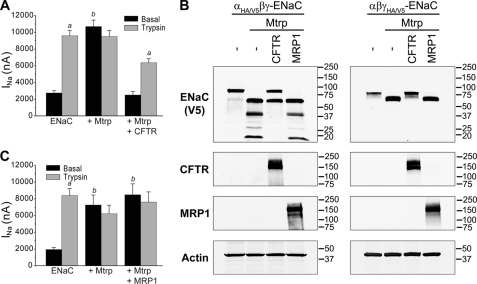
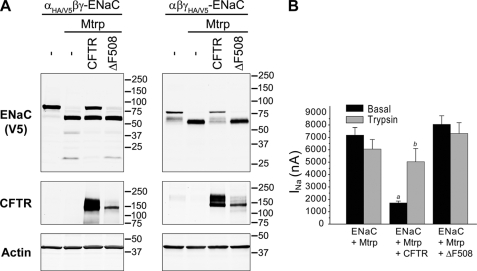
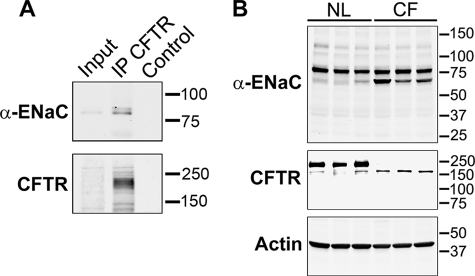
Cystic fibrosis (CF) is caused by mutations in the CF transmembrane conductance regulator (CFTR) that prevent its proper folding and trafficking to the apical membrane of epithelial cells. Absence of cAMP-mediated Cl(-) secretion in CF airways causes poorly hydrated airway surfaces in CF patients, and this condition is exacerbated by excessive Na(+) absorption. The mechanistic link between missing CFTR and increased Na(+) absorption in airway epithelia has remained elusive, although substantial evidence implicates hyperactivity of the epithelial Na(+) channel (ENaC). ENaC is known to be activated by selective endoproteolysis of the extracellular domains of its α- and γ-subunits, and it was recently reported that ENaC and CFTR physically associate in mammalian cells. We confirmed this interaction in oocytes by co-immunoprecipitation and found that ENaC associated with wild-type CFTR was protected from proteolytic cleavage and stimulation of open probability. In contrast, ΔF508 CFTR, the most common mutant protein in CF patients, failed to protect ENaC from proteolytic cleavage and stimulation. In normal airway epithelial cells, ENaC was contained in the anti-CFTR immunoprecipitate. In CF airway epithelial cultures, the proportion of full-length to total α-ENaC protein signal was consistently reduced compared with normal cultures. Our results identify limiting proteolytic cleavage of ENaC as a mechanism by which CFTR down-regulates Na(+) absorption.
Figures
Comment in
-
CFTR's proteolysis protection. The cystic fibrosis transmembrane conductance regulator impedes proteolytic stimulation of the epithelial Na+ channel.J Biol Chem. 2010 Oct 15;285(42):e99974. doi: 10.1074/jbc.p110.155259. J Biol Chem. 2010. PMID: 20954295 Free PMC article. No abstract available.
References
-
- Matsui H., Grubb B. R., Tarran R., Randell S. H., Gatzy J. T., Davis C. W., Boucher R. C. (1998) Cell 95, 1005–1015 - PubMed
-
- Boucher R. C. (2003) Pflugers Arch. 445, 495–498 - PubMed
-
- Stutts M. J., Canessa C. M., Olsen J. C., Hamrick M., Cohn J. A., Rossier B. C., Boucher R. C. (1995) Science 269, 847–850 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
