

The melanoma-associated transmembrane glycoprotein Gpnmb controls trafficking of cellular debris for degradation and is essential for tissue repair
- PMID: 20709912
- PMCID: PMC2992370
- DOI: 10.1096/fj.10-154757
The melanoma-associated transmembrane glycoprotein Gpnmb controls trafficking of cellular debris for degradation and is essential for tissue repair
Abstract
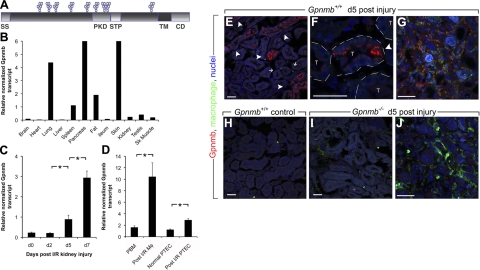
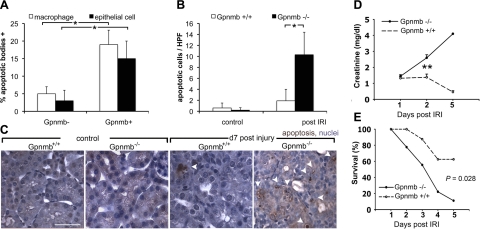
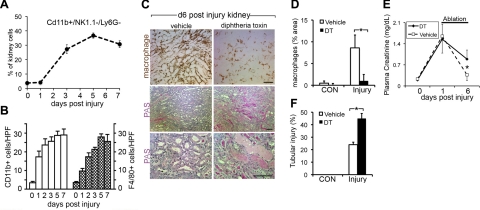
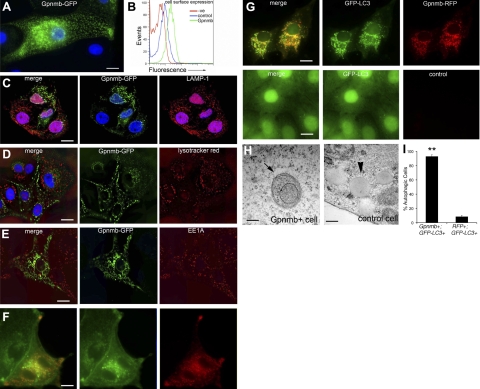
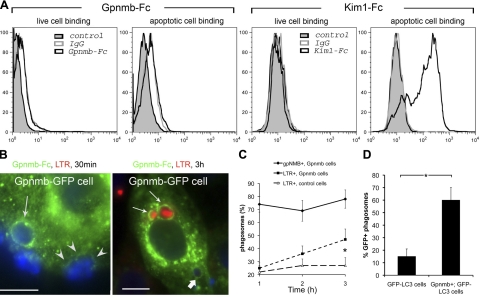
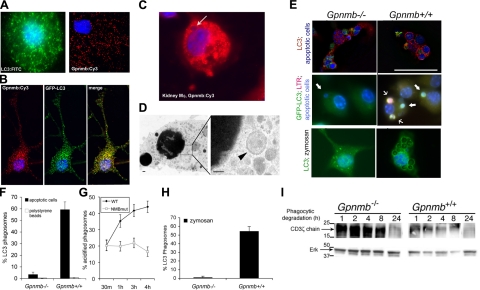
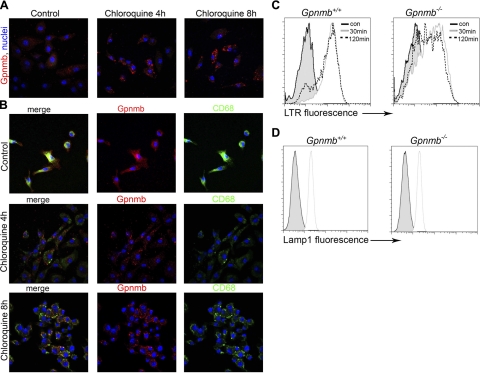
Kidney damage due to injury rarely resolves completely, and there are currently no therapies capable of promoting repair. In addition to understanding mechanisms by which tissues are damaged, illuminating mechanisms of repair and regeneration is also of great importance. Here we show that the melanoma-associated, transmembrane glycoprotein, Gpnmb, is up-regulated 15-fold following ischemic damage in kidney tissue and by more than 10-fold in macrophages and 3-fold in surviving epithelial cells. Gpnmb-expressing macrophages and epithelial cells were found to contain apoptotic bodies at 3 times the rate of nonexpressing cells. Either mutation of Gpnmb or ablation of inflammatory macrophages prevents normal repair of the kidney. Significantly, the kidneys from postischemic Gpnmb mutant mice exhibited a 5-fold increase in apoptotic cellular debris compared to wild-type mice. These mice also experienced an 85% increase in mortality following bilateral ischemic kidney. Finally, we demonstrate that Gpnmb is a phagocytic protein that is necessary for recruitment of the autophagy protein LC3 to the phagosome where these proteins are colocalized and for lysosomal fusion with the phagosome and hence bulk degradation of their content. Therefore, Gpnmb is a novel prorepair gene that is necessary for crosstalk between the macroautophagic degradation pathway and phagocytosis.
Figures
References
-
- Duffield J. S., Hong S., Vaidya V. S., Lu Y., Fredman G., Serhan C. N., Bonventre J. V. (2006) Resolvin D series and protectin D1 mitigate acute kidney injury. J. Immunol. 177, 5902–5911 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
