

FSH dystrophy and a subtelomeric 4q haplotype: a new assay and associations with disease
- PMID: 20710047
- PMCID: PMC3043595
- DOI: 10.1136/jmg.2009.076703
FSH dystrophy and a subtelomeric 4q haplotype: a new assay and associations with disease
Abstract
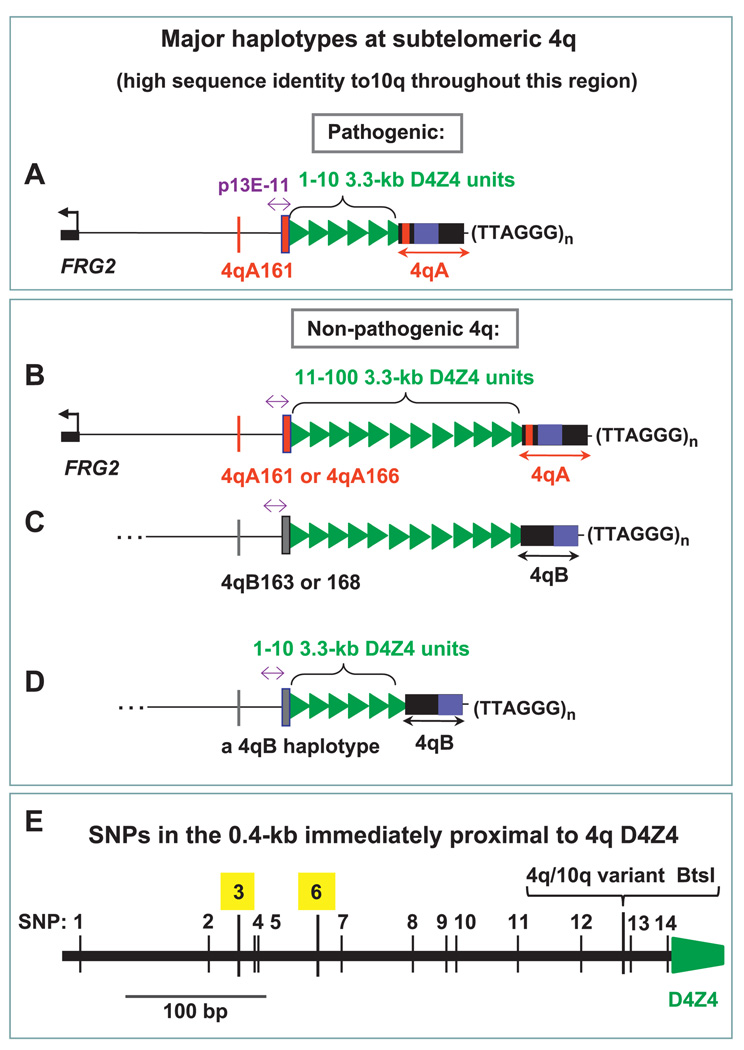
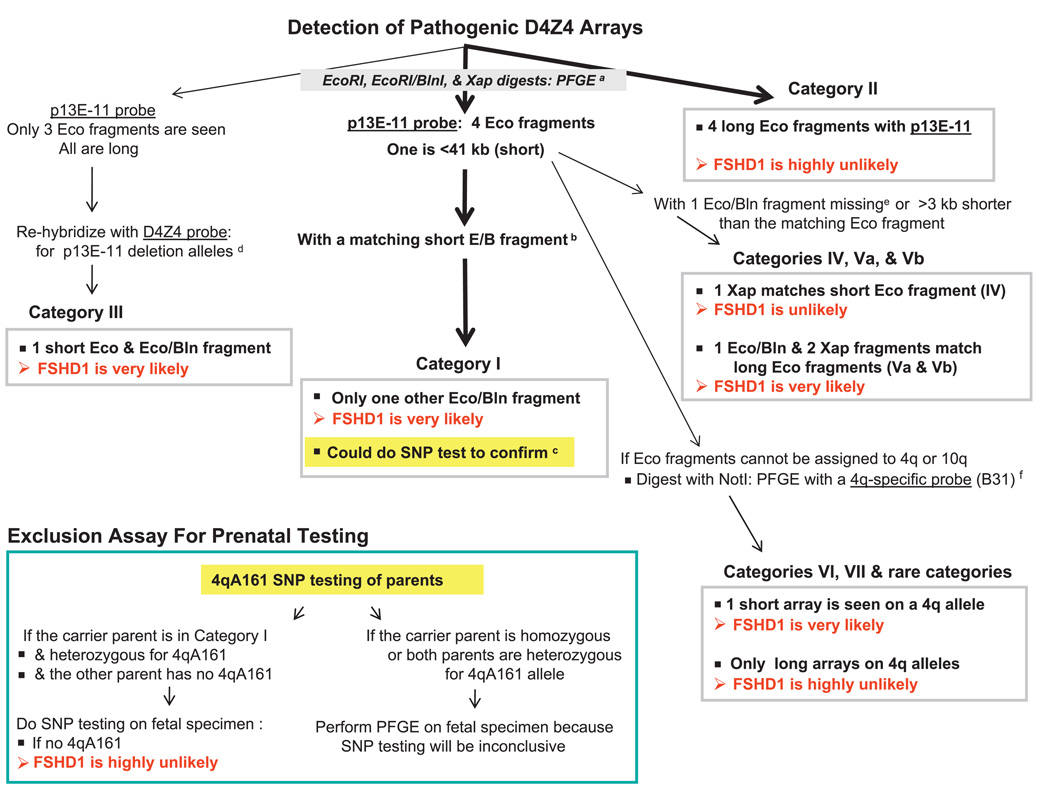
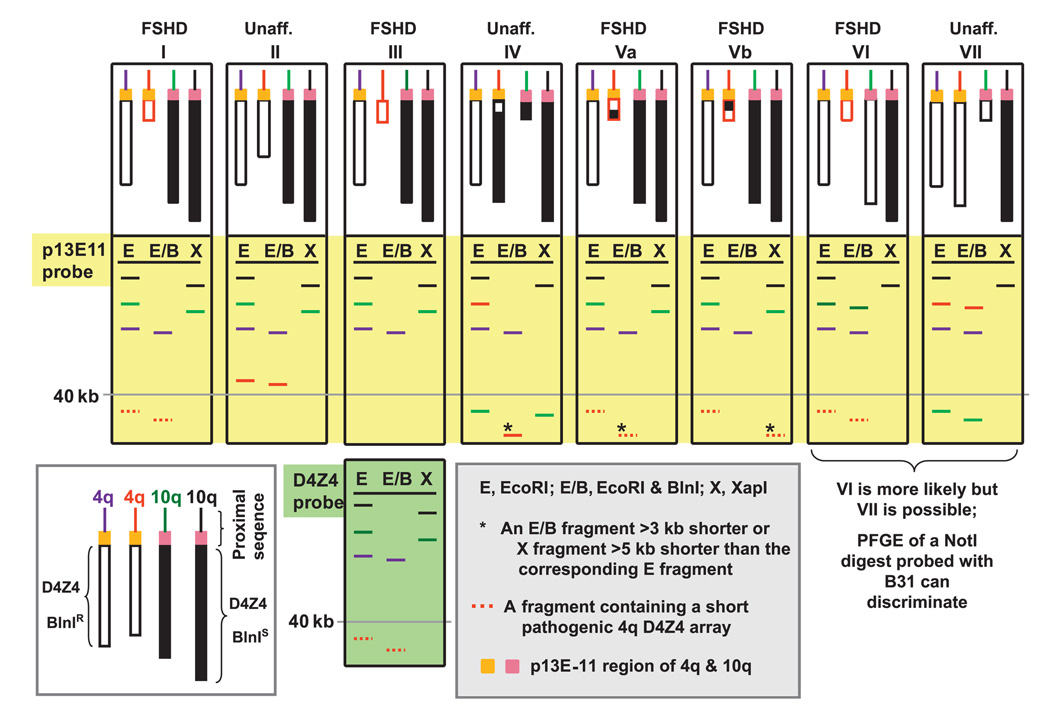
Background: Facioscapulohumeral muscular dystrophy (FSHD) is an autosomal dominant disease associated with contraction of arrays of tandem 3.3-kb units (D4Z4) on subtelomeric 4q. Disease-linked arrays usually have fewer than 11 repeat units. Equally short D4Z4 arrays at subtelomeric 10q are not linked to FSHD. The newly described 4qA161 haplotype, which is more prevalent in pathogenic 4q alleles, involves sequences in and near D4Z4.
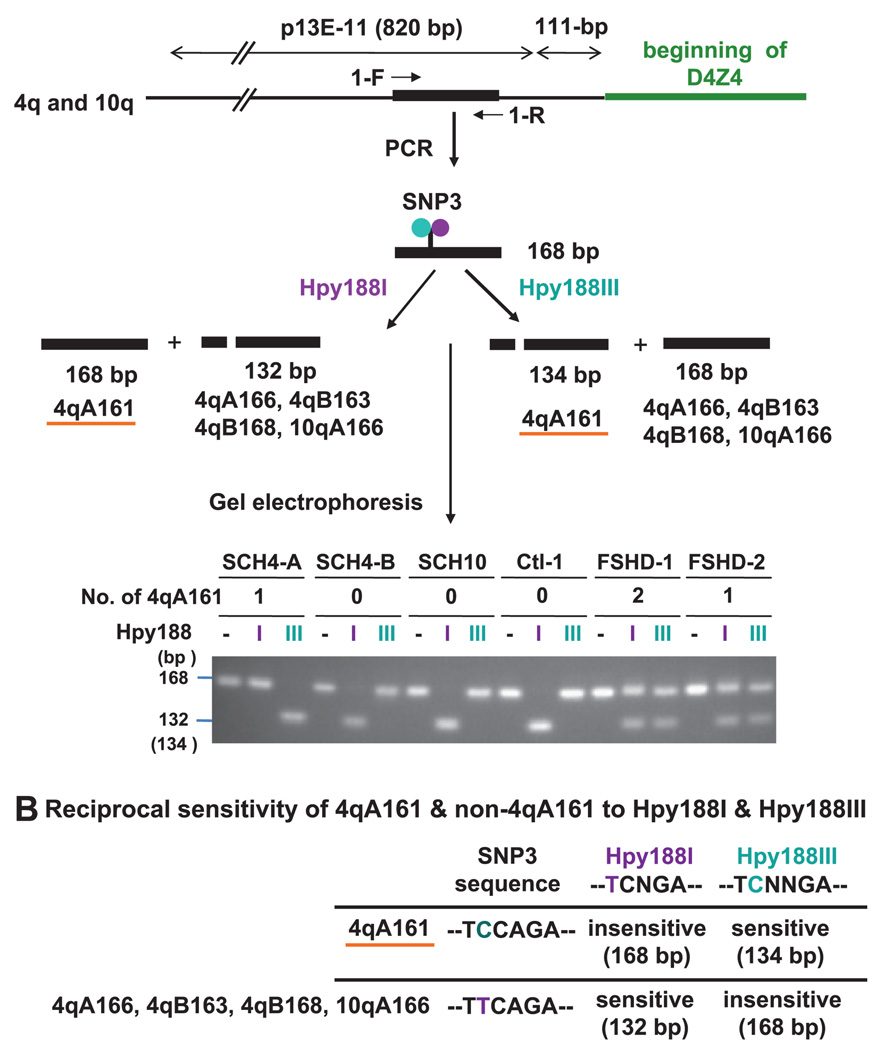
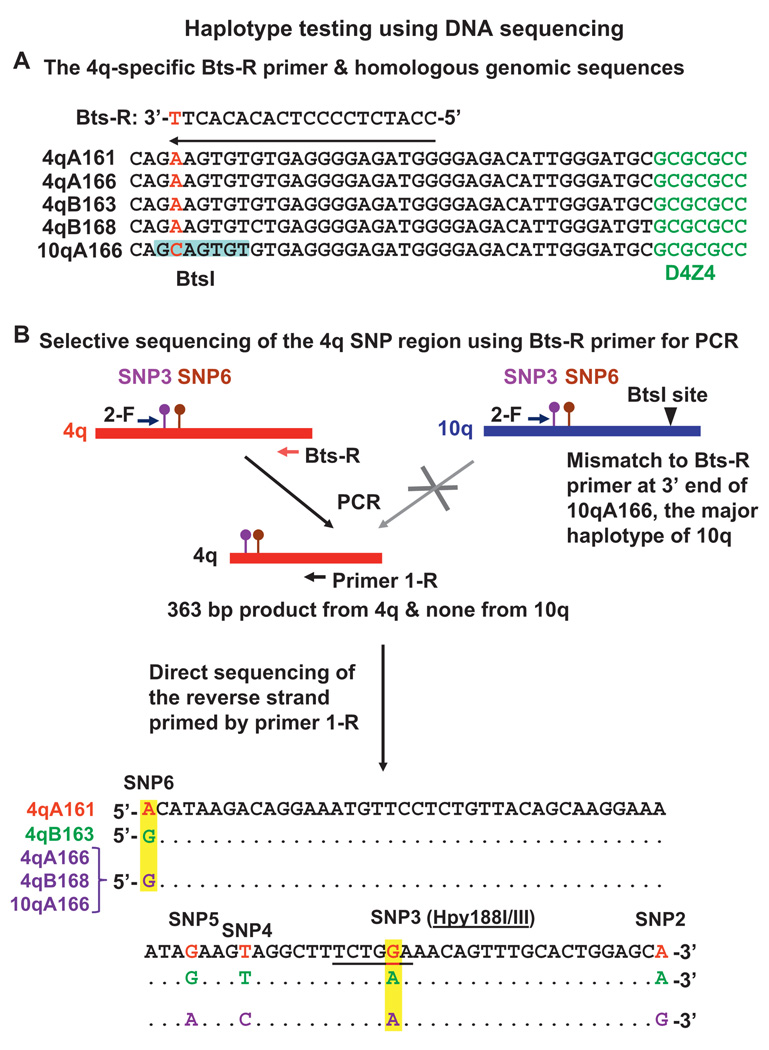
Methods: We developed two new assays for 4qA161, which are based upon direct sequencing of PCR products or detecting restriction fragment length polymorphisms. They were used to analyse single nucleotide polymorphisms (SNPs) indicative of 4q161 alleles.
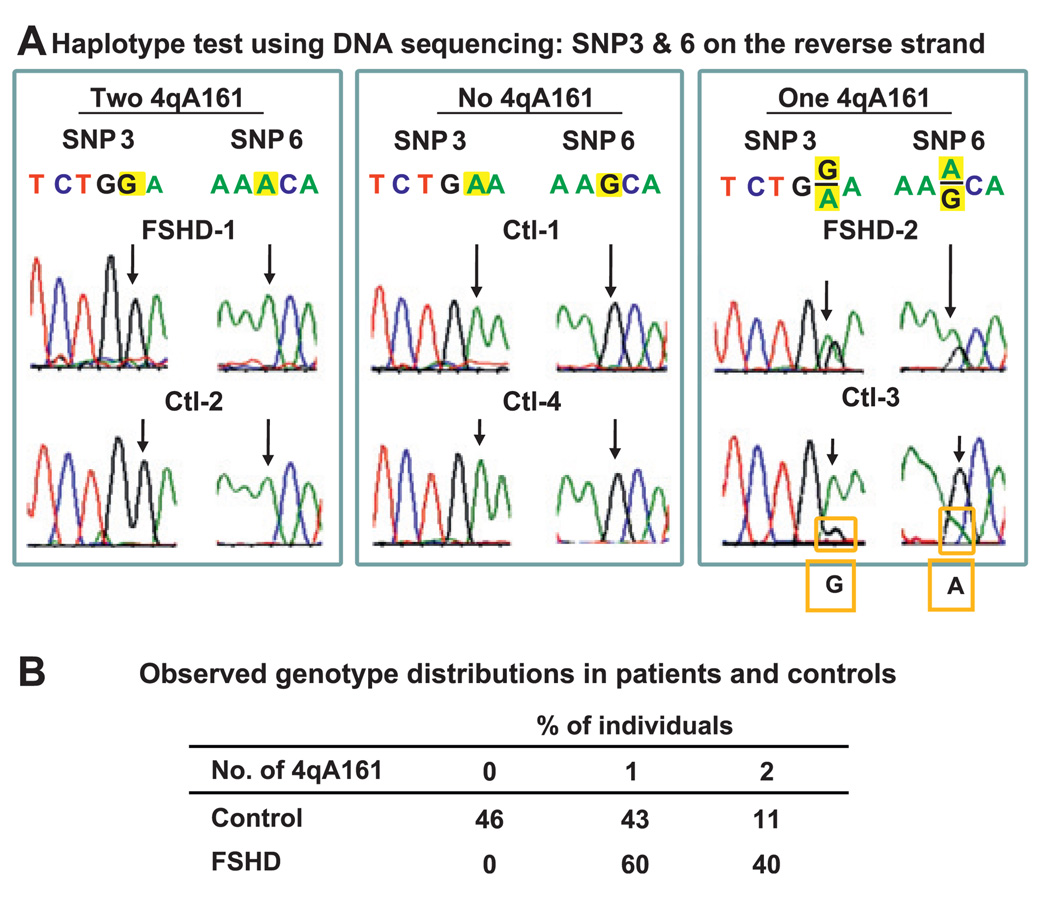
Results: All (35/35) FSHD patients had one or two 4qA161 alleles (60% or 40%, respectively). In contrast, 46% (21/46) of control individuals had no 4qA161 allele (p<10(-4)), and 26% had homozygous 4qB163 alleles.
Conclusions: Our results from a heterogeneous population are consistent with the previously described association of the 4qA161 haplotype with FSHD, but a causal association with pathogenesis is uncertain. In addition, we found that haplotype analysis is complicated by the presence of minor 10q alleles. Nonetheless, our sequencing assay for the 4qA161allele can enhance molecular diagnosis of FSHD, including prenatal diagnosis, and is simpler to perform than the previously described assay.
Figures
References
-
- Lunt P. Facioscapulohumeral Muscular Dystrophy: Diagnostic and Molecular Aspects. In: Deymeer F, editor. Neuromuscular Diseases: From Basic Mechanisms to Clinical Management. Vol 18: Monogr. Clin. Neurosci. Basel: Karger; 2000. pp. 44–60.
-
- Pandya S, King WM, Tawil R. Facioscapulohumeral dystrophy. Phys Ther. 2008;88:105–113. - PubMed
-
- van Deutekom JC, Wijmenga C, van Tienhoven EA, Gruter AM, Hewitt JE, Padberg GW, van Ommen GJ, Hofker MH, Frants RR. FSHD associated DNA rearrangements are due to deletions of integral copies of a 3.2 kb tandemly repeated unit. Hum Mol Genet. 1993;2:2037–2042. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources