

Telomere disruption results in non-random formation of de novo dicentric chromosomes involving acrocentric human chromosomes
- PMID: 20711355
- PMCID: PMC2920838
- DOI: 10.1371/journal.pgen.1001061
Telomere disruption results in non-random formation of de novo dicentric chromosomes involving acrocentric human chromosomes
Abstract
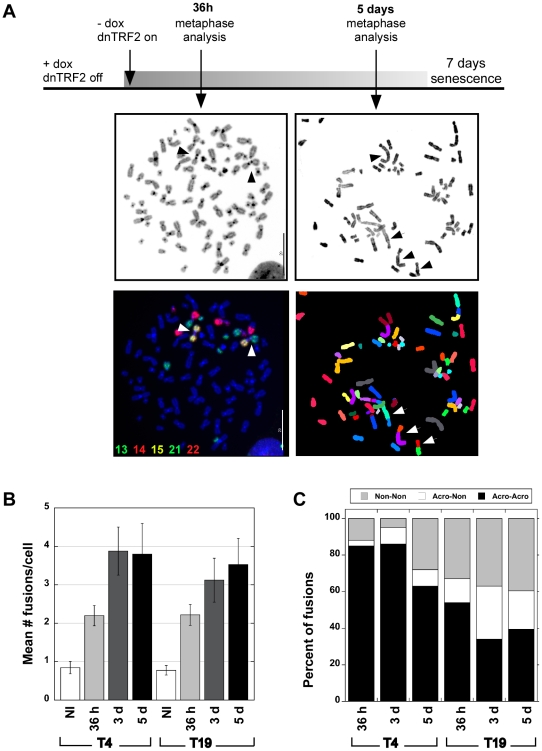
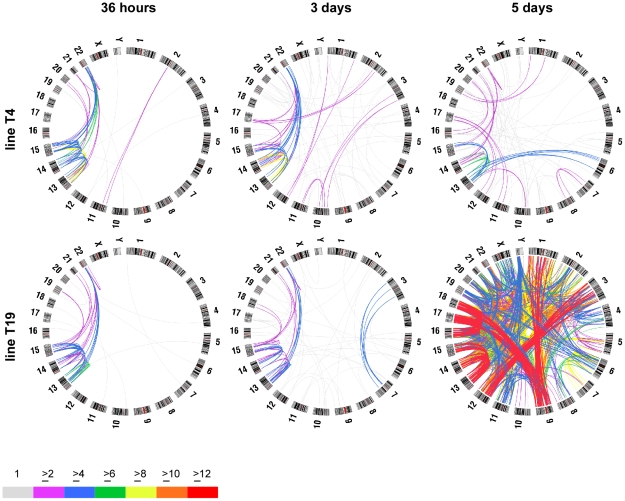
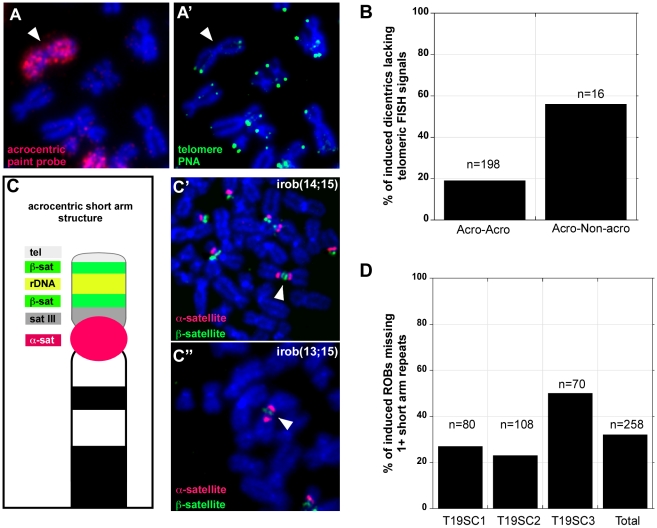
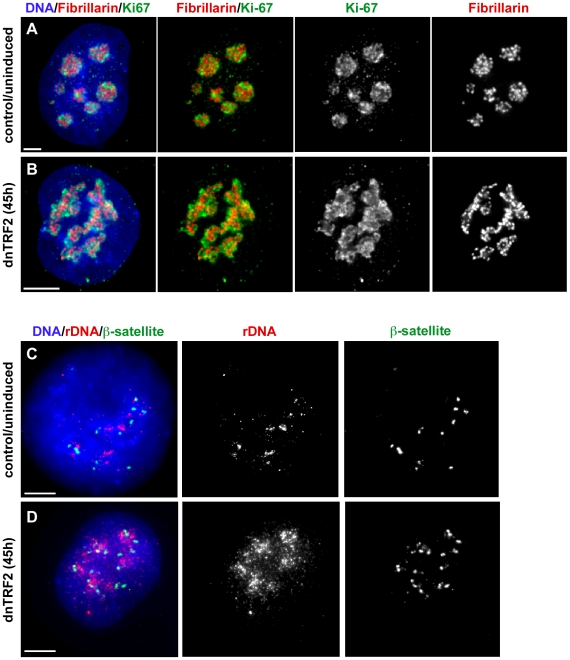
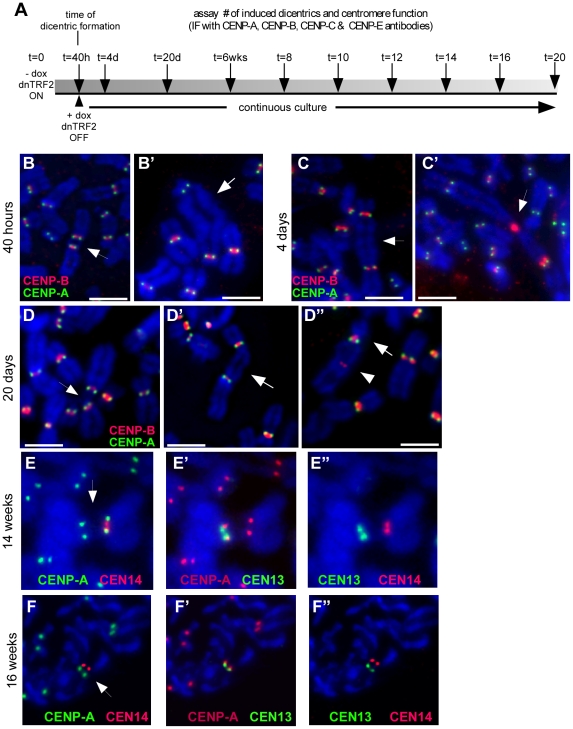
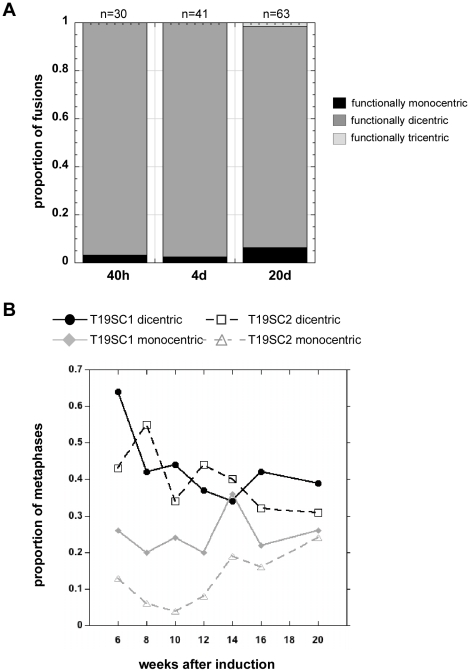
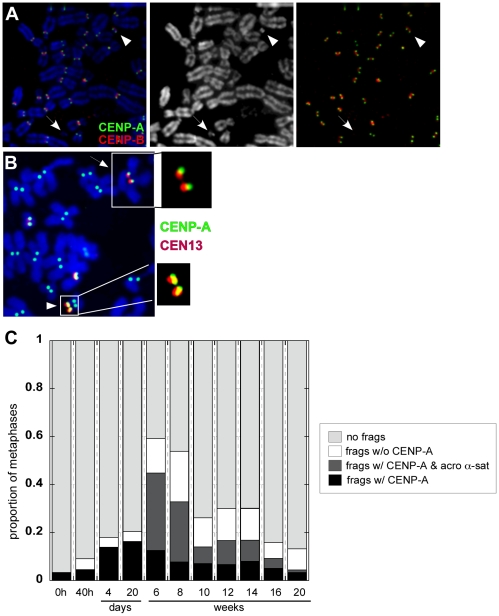
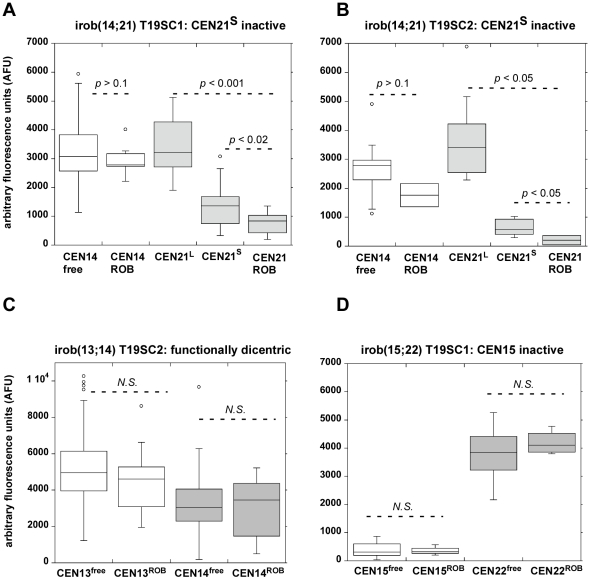
Genome rearrangement often produces chromosomes with two centromeres (dicentrics) that are inherently unstable because of bridge formation and breakage during cell division. However, mammalian dicentrics, and particularly those in humans, can be quite stable, usually because one centromere is functionally silenced. Molecular mechanisms of centromere inactivation are poorly understood since there are few systems to experimentally create dicentric human chromosomes. Here, we describe a human cell culture model that enriches for de novo dicentrics. We demonstrate that transient disruption of human telomere structure non-randomly produces dicentric fusions involving acrocentric chromosomes. The induced dicentrics vary in structure near fusion breakpoints and like naturally-occurring dicentrics, exhibit various inter-centromeric distances. Many functional dicentrics persist for months after formation. Even those with distantly spaced centromeres remain functionally dicentric for 20 cell generations. Other dicentrics within the population reflect centromere inactivation. In some cases, centromere inactivation occurs by an apparently epigenetic mechanism. In other dicentrics, the size of the alpha-satellite DNA array associated with CENP-A is reduced compared to the same array before dicentric formation. Extra-chromosomal fragments that contained CENP-A often appear in the same cells as dicentrics. Some of these fragments are derived from the same alpha-satellite DNA array as inactivated centromeres. Our results indicate that dicentric human chromosomes undergo alternative fates after formation. Many retain two active centromeres and are stable through multiple cell divisions. Others undergo centromere inactivation. This event occurs within a broad temporal window and can involve deletion of chromatin that marks the locus as a site for CENP-A maintenance/replenishment.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Hanahan D, Weinberg R. The hallmarks of cancer. Cell. 2000;100:57–70. - PubMed
-
- Mitelman F, Mertens F, Johansson B. A breakpoint map of recurrent chromosomal rearrangements in human neoplasia. Nat Genet. 1997;15:417–474. - PubMed
-
- Mitelman F. Recurrent chromosome aberrations in cancer. Mutation Research/Review in Mutation Research. 2000;462:247–253. - PubMed
-
- Hamerton J, Canning N, Ray M, Smith S. A cytogenetic survey of 14,069 newborn infants. I. Incidence of chromosome abnormalities. Clin Genet. 1975;8:223–243. - PubMed
-
- Robertson W. Chromosome studies. I. Taxonomic relationships shown in the chromosomes of Tettigidae and Acrididae. V-shaped chromosomes and their significant in Acrididae, Locustidae and Gryllidae: chromosome and variation. J Morph. 1916;27:179–331.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
