

A variable region in GP4 of European-type porcine reproductive and respiratory syndrome virus induces neutralizing antibodies against homologous but not heterologous virus strains
- PMID: 20712485
- PMCID: PMC2928701
- DOI: 10.1089/vim.2010.0025
A variable region in GP4 of European-type porcine reproductive and respiratory syndrome virus induces neutralizing antibodies against homologous but not heterologous virus strains
Abstract
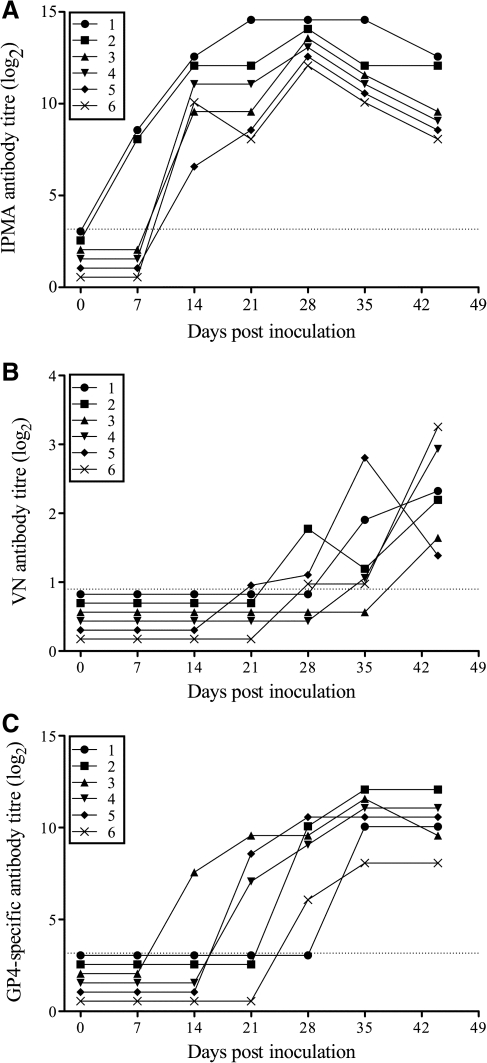
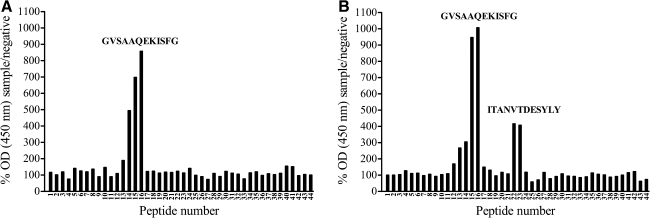
Porcine reproductive and respiratory syndrome virus (PRRSV) can induce severe reproductive failure in sows, and is involved in the porcine respiratory disease complex. The glycoprotein GP4 of the European prototype PRRSV strain Lelystad virus (LV) contains a linear neutralizing epitope that is located in a highly variable region. The current study aimed to evaluate the antibody response against this and other epitopes on GP4 to infection of pigs with European-type PRRSV. It was shown that three virus strains, differing in the region that corresponds to the neutralizing epitope on GP4 of LV, strongly induce antibodies against this area. Antibodies against the epitopes of the different virus strains were purified from polyclonal swine sera, and used in virus-neutralization tests on primary alveolar macrophages. This revealed that antibodies against the variable region in GP4 of different virus strains are able to neutralize infection with homologous but not heterologous virus strains.
Figures

Comment in
-
A focus on humoral immunity.Viral Immunol. 2010 Aug;23(4):341. doi: 10.1089/vim.2010.ed23.4. Viral Immunol. 2010. PMID: 20712477 No abstract available.
References
-
- Allende R. Lewis TL. Lu Z, et al. North American and European porcine reproductive and respiratory syndrome viruses differ in non-structural protein coding regions. J Gen Virol. 1999;80(Pt 2):307–315. - PubMed
-
- Christianson WT. Collins JE. Benfield DA, et al. Experimental reproduction of swine infertility and respiratory syndrome in pregnant sows. Am J Vet Res. 1992;53:485–488. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
