

Magnesium-dependent interaction of PKR with adenovirus VAI
- PMID: 20713064
- PMCID: PMC2980848
- DOI: 10.1016/j.jmb.2010.08.015
Magnesium-dependent interaction of PKR with adenovirus VAI
Abstract
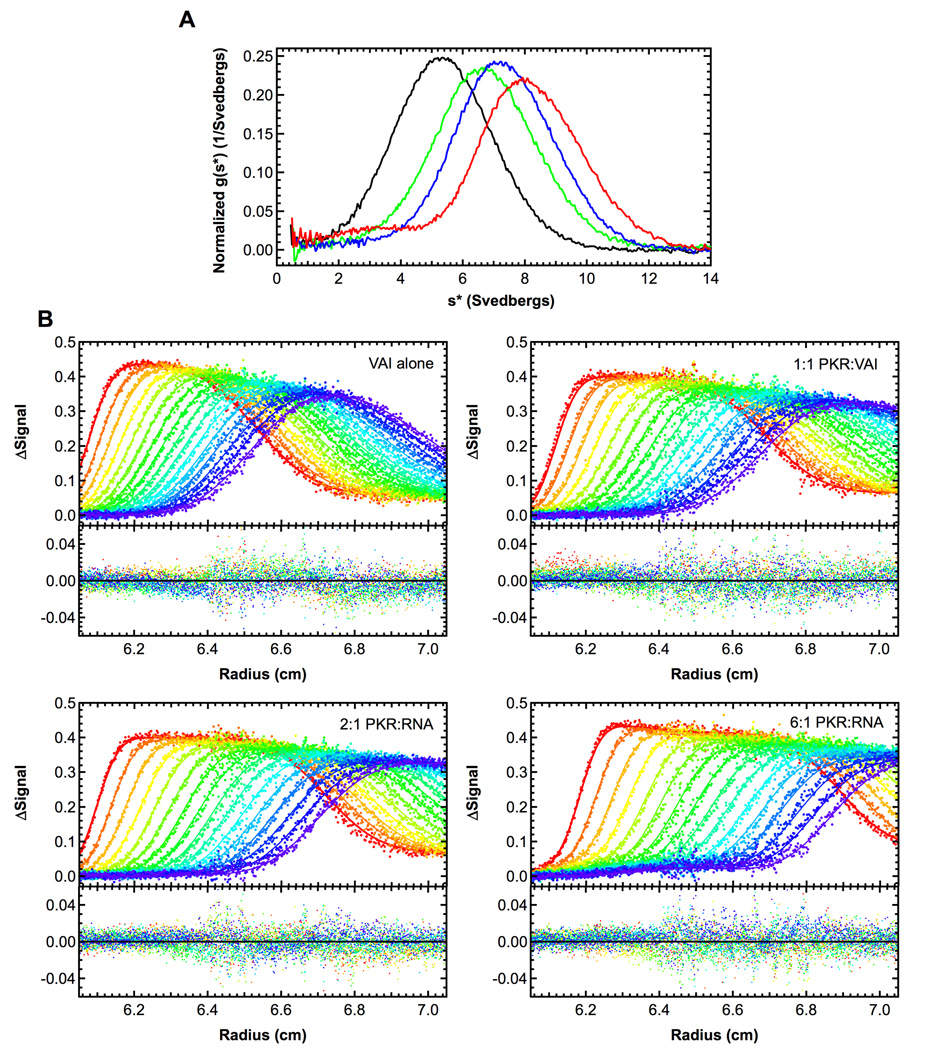
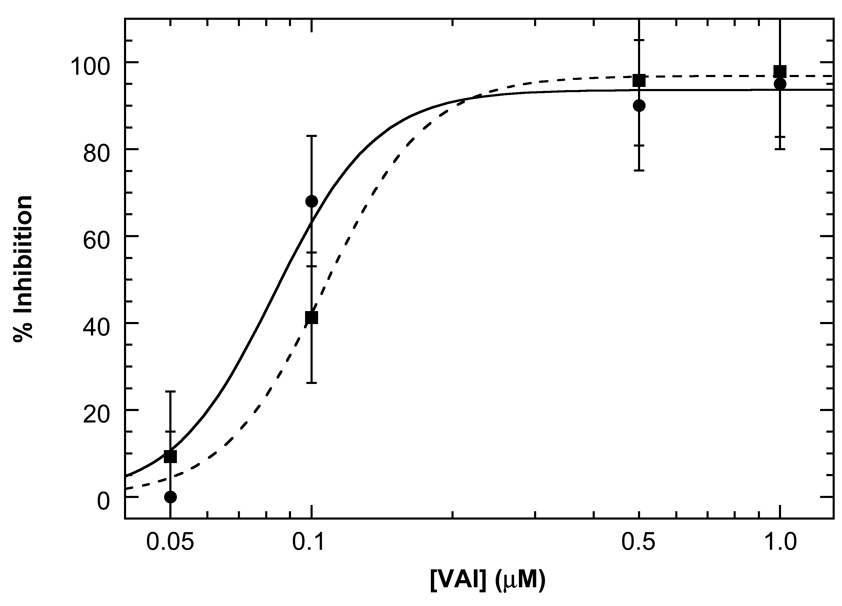
Protein kinase R (PKR) is an interferon-induced kinase that plays a pivotal role in the innate immunity pathway for defense against viral infection. PKR is activated to undergo autophosphorylation upon binding to RNAs that contain duplex regions. Activated PKR phosphorylates the α-subunit of eukaryotic initiation factor 2, thereby inhibiting protein synthesis in virus-infected cells. Viruses have evolved diverse PKR-inhibitory strategies to evade the antiviral response. Adenovirus encodes virus-associated RNA I (VAI), a highly structured RNA inhibitor that binds PKR but fails to activate. We have characterized the stoichiometry and affinity of PKR binding to define the mechanism of PKR inhibition by VAI. Sedimentation velocity and isothermal titration calorimetry measurements indicate that PKR interactions with VAI are modulated by Mg(2+). Two PKR monomers bind in the absence of Mg(2+), but a single monomer binds in the presence of divalent ion. Known RNA activators of PKR are capable of binding multiple PKR monomers to allow the kinase domains to come into close proximity and thus enhance dimerization. We propose that VAI acts as an inhibitor of PKR because it binds and sequesters a single PKR in the presence of divalent cation.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures


References
-
- Toth AM, Zhang P, Das S, George CX, Samuel CE. Interferon action and the double-stranded RNA-dependent enzymes ADAR1 adenosine deaminase and PKR protein kinase. Prog. Nucleic Acid Res. Mol. Biol. 2006;81:369–434. - PubMed
-
- Tian B, Bevilacqua PC, Diegelman-Parente A, Mathews MB. The double-stranded RNA binding motif: Interference and much more. Nature Rev. Mol. Cell Biol. 2004;5:1013–1023. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
