

RssAB-FlhDC-ShlBA as a major pathogenesis pathway in Serratia marcescens
- PMID: 20713626
- PMCID: PMC2976324
- DOI: 10.1128/IAI.00661-10
RssAB-FlhDC-ShlBA as a major pathogenesis pathway in Serratia marcescens
Abstract
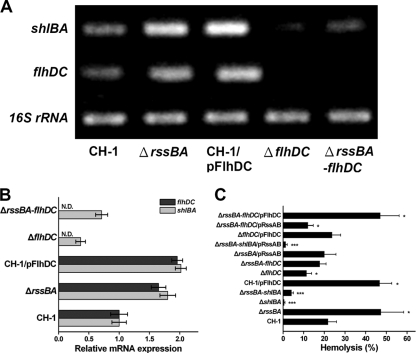
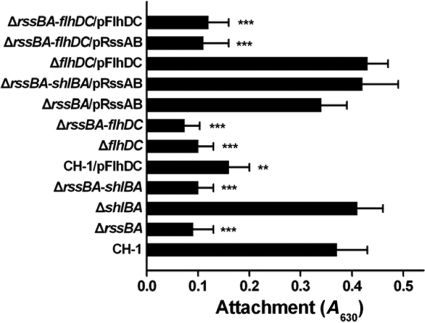
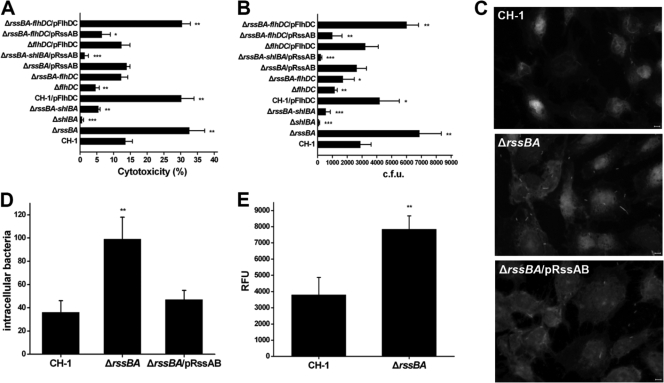
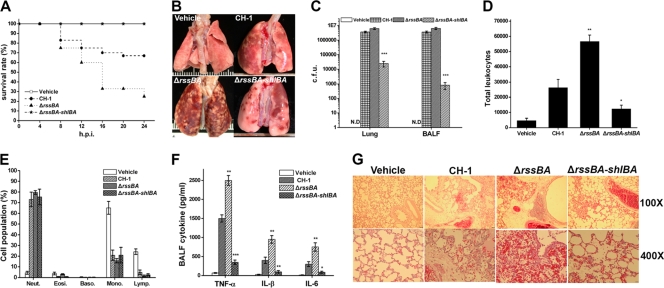
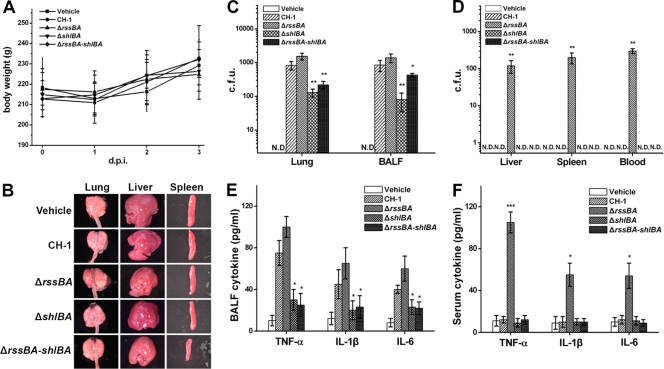
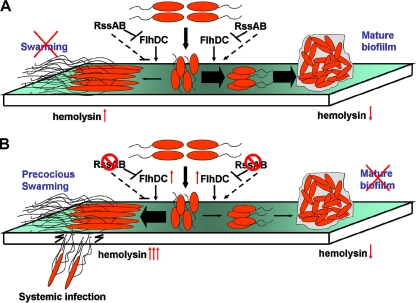
Serratia marcescens has long been recognized as an important opportunistic pathogen, but the underlying pathogenesis mechanism is not completely clear. Here, we report a key pathogenesis pathway in S. marcescens comprising the RssAB two-component system and its downstream elements, FlhDC and the dominant virulence factor hemolysin ShlBA. Expression of shlBA is under the positive control of FlhDC, which is repressed by RssAB signaling. At 37°C, functional RssAB inhibits swarming, represses hemolysin production, and promotes S. marcescens biofilm formation. In comparison, when rssBA is deleted, S. marcescens displays aberrant multicellularity favoring motile swarming with unbridled hemolysin production. Cellular and animal infection models further demonstrate that loss of rssBA transforms this opportunistic pathogen into hypervirulent phenotypes, leading to extensive inflammatory responses coupled with destructive and systemic infection. Hemolysin production is essential in this context. Collectively, a major virulence regulatory pathway is identified in S. marcescens.
Figures
References
-
- Buffet-Bataillon, S., V. Rabier, P. Betremieux, A. Beuchee, M. Bauer, P. Pladys, E. Le Gall, M. Cormier, and A. Jolivet-Gougeon. 2009. Outbreak of Serratia marcescens in a neonatal intensive care unit: contaminated unmedicated liquid soap and risk factors. J. Hosp. Infect. 72:17-22. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
