

Gelatinase contributes to the pathogenesis of endocarditis caused by Enterococcus faecalis
- PMID: 20713628
- PMCID: PMC2976315
- DOI: 10.1128/IAI.01118-09
Gelatinase contributes to the pathogenesis of endocarditis caused by Enterococcus faecalis
Abstract
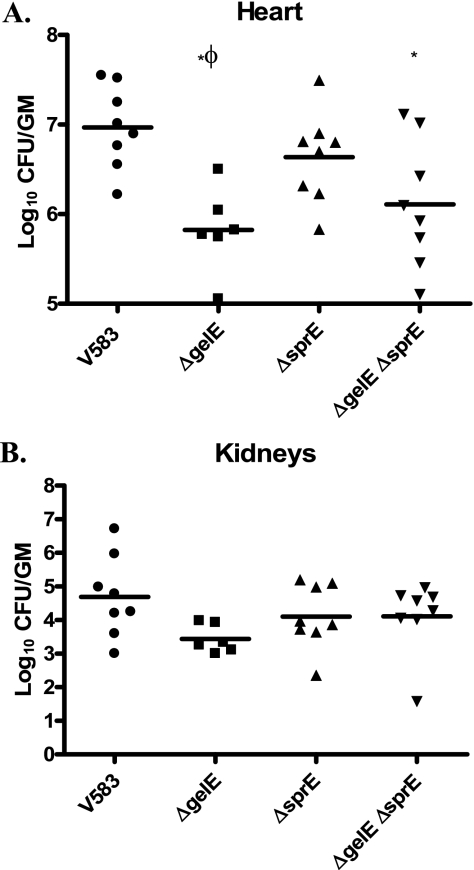
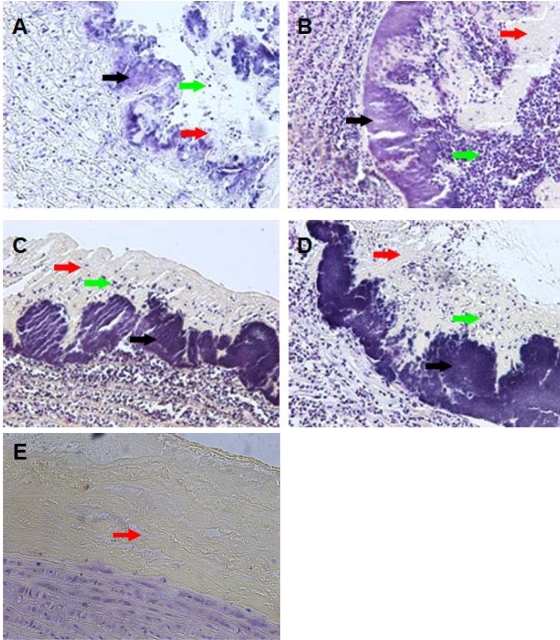
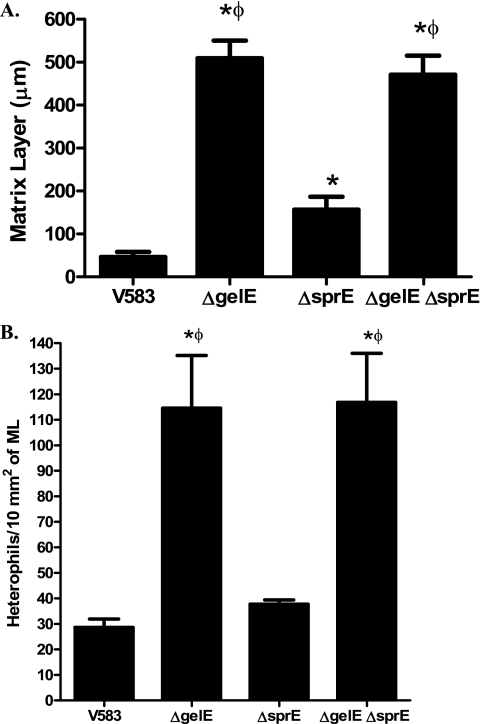
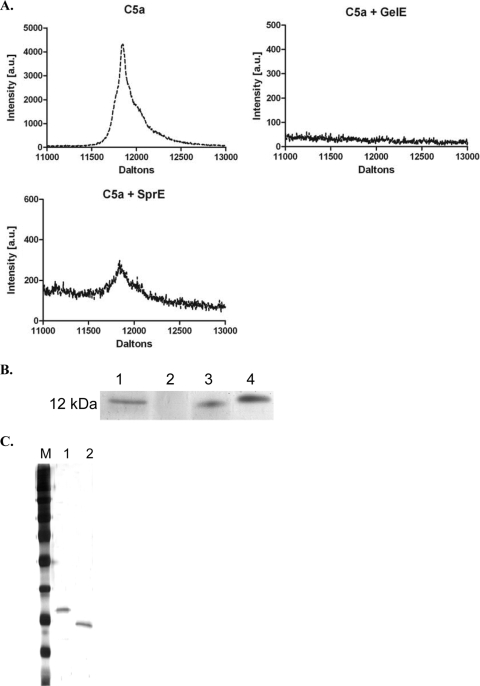
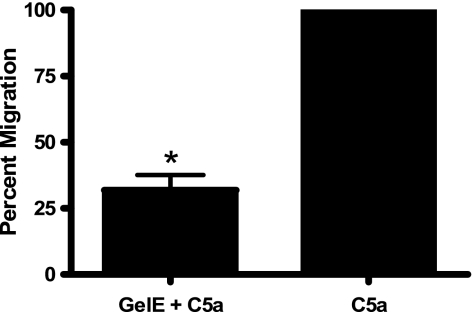
The Gram-positive pathogen Enterococcus faecalis is a leading agent of nosocomial infections, including urinary tract infections, surgical site infections, and bacteremia. Among the infections caused by E. faecalis, endocarditis remains a serious clinical manifestation and unique in that it is commonly acquired in a community setting. Infective endocarditis is a complex disease, with many host and microbial components contributing to the formation of bacterial biofilm-like vegetations on the aortic valve and adjacent areas within the heart. In the current study, we compared the pathogenic potential of the vancomycin-resistant E. faecalis V583 and three isogenic protease mutants (ΔgelE, ΔsprE, and ΔgelE ΔsprE mutants) in a rabbit model of enterococcal endocarditis. The bacterial burdens displayed by GelE(-) mutants (ΔgelE and ΔgelE ΔsprE mutants) in the heart were significantly lower than those of V583 or the SprE(-) mutant. Vegetations on the aortic valve infected with GelE(-) mutants (ΔgelE and ΔgelE ΔsprE mutants) also showed a significant increase in deposition of fibrinous matrix layer and increased chemotaxis of inflammatory cells. In support of a role for proteolytic modulation of the immune response to E. faecalis, we also demonstrate that GelE can cleave the anaphylatoxin complement C5a and that this proteolysis leads to decreased neutrophil migration in vitro. In vivo, a decreased heterophil (neutrophil-like cell) migration was observed at tissue sites infected with GelE-producing strains but not at those infected with SprE-producing strains. Taken together, these observations suggest that of the two enterococcal proteases, gelatinase is the principal mediator of pathogenesis in endocarditis.
Figures
References
-
- DiScipio, R. G., P. J. Daffern, M. A. Jagels, D. H. Broide, and P. Sriramarao. 1999. A comparison of C3a and C5a-mediated stable adhesion of rolling eosinophils in postcapillary venules and transendothelial migration in vitro and in vivo. J. Immunol. 162:1127-1136. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
