

Stabilization of neurotoxic Alzheimer amyloid-beta oligomers by protein engineering
- PMID: 20713699
- PMCID: PMC2932621
- DOI: 10.1073/pnas.1001740107
Stabilization of neurotoxic Alzheimer amyloid-beta oligomers by protein engineering
Abstract
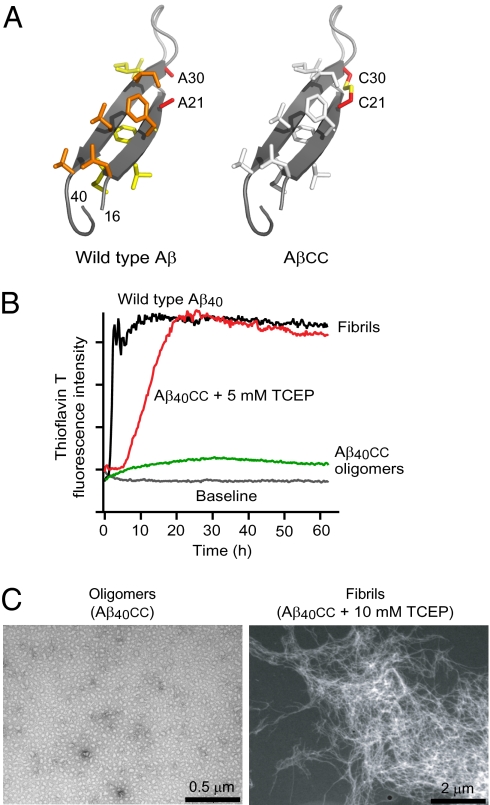
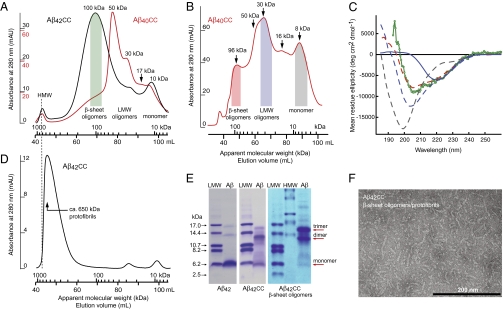
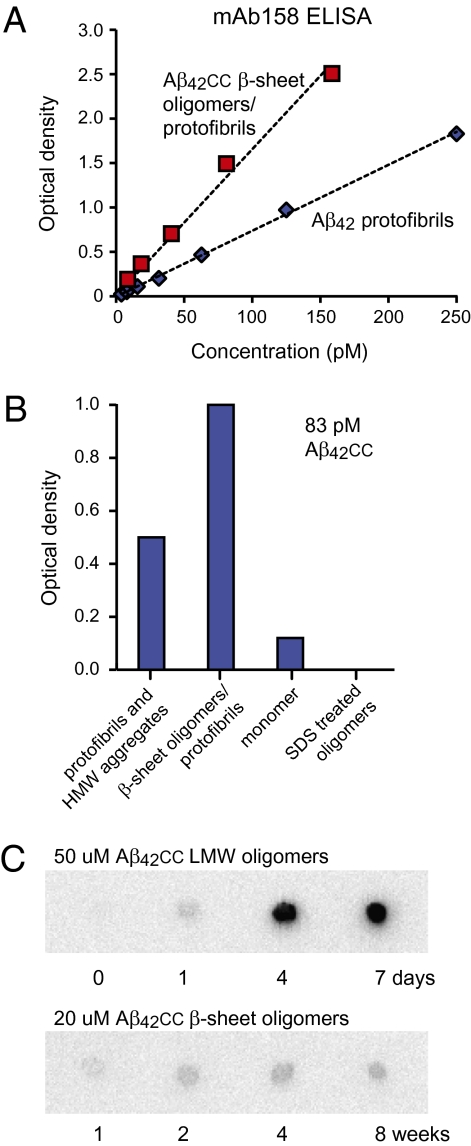
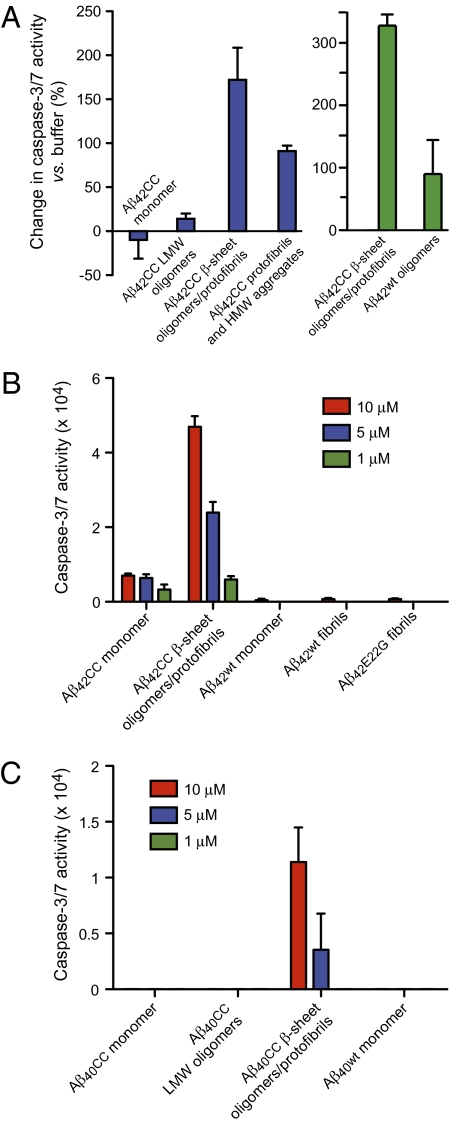
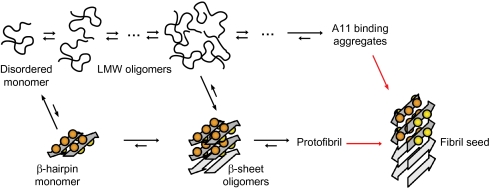
Soluble oligomeric aggregates of the amyloid-beta peptide (Abeta) have been implicated in the pathogenesis of Alzheimer's disease (AD). Although the conformation adopted by Abeta within these aggregates is not known, a beta-hairpin conformation is known to be accessible to monomeric Abeta. Here we show that this beta-hairpin is a building block of toxic Abeta oligomers by engineering a double-cysteine mutant (called Abetacc) in which the beta-hairpin is stabilized by an intramolecular disulfide bond. Abeta(40)cc and Abeta(42)cc both spontaneously form stable oligomeric species with distinct molecular weights and secondary-structure content, but both are unable to convert into amyloid fibrils. Biochemical and biophysical experiments and assays with conformation-specific antibodies used to detect Abeta aggregates in vivo indicate that the wild-type oligomer structure is preserved and stabilized in Abetacc oligomers. Stable oligomers are expected to become highly toxic and, accordingly, we find that beta-sheet-containing Abeta(42)cc oligomers or protofibrillar species formed by these oligomers are 50 times more potent inducers of neuronal apoptosis than amyloid fibrils or samples of monomeric wild-type Abeta(42), in which toxic aggregates are only transiently formed. The possibility of obtaining completely stable and physiologically relevant neurotoxic Abeta oligomer preparations will facilitate studies of their structure and role in the pathogenesis of AD. For example, here we show how kinetic partitioning into different aggregation pathways can explain why Abeta(42) is more toxic than the shorter Abeta(40), and why certain inherited mutations are linked to protofibril formation and early-onset AD.
Conflict of interest statement
Conflict of interest statement: A.S. and T.H. are shareholders of MIVAC Development AB, Gothenburg, Sweden.
Figures
References
-
- Chiti F, Dobson CM. Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem. 2006;75:333–366. - PubMed
-
- Lesné S, et al. A specific amyloid-β protein assembly in the brain impairs memory. Nature. 2006;440:352–357. - PubMed
-
- De Felice FG, et al. Abeta oligomers induce neuronal oxidative stress through an N-methyl-D-aspartate receptor-dependent mechanism that is blocked by the Alzheimer drug memantine. J Biol Chem. 2007;282:11590–11601. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
