

Tip-DC development during parasitic infection is regulated by IL-10 and requires CCL2/CCR2, IFN-gamma and MyD88 signaling
- PMID: 20714353
- PMCID: PMC2920868
- DOI: 10.1371/journal.ppat.1001045
Tip-DC development during parasitic infection is regulated by IL-10 and requires CCL2/CCR2, IFN-gamma and MyD88 signaling
Abstract
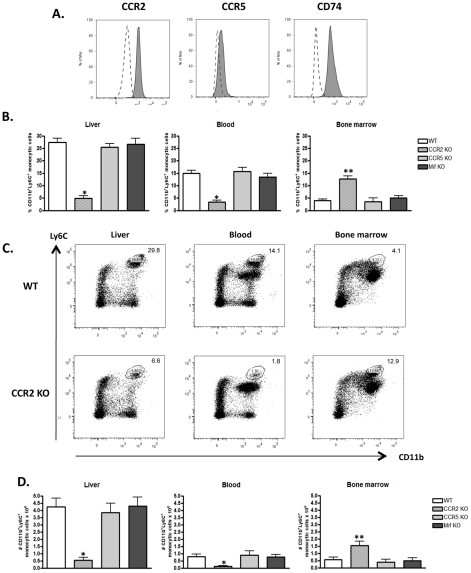
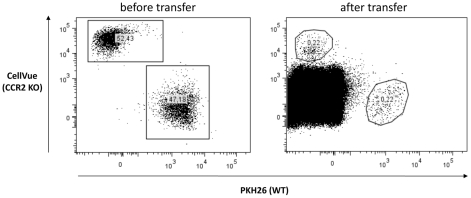
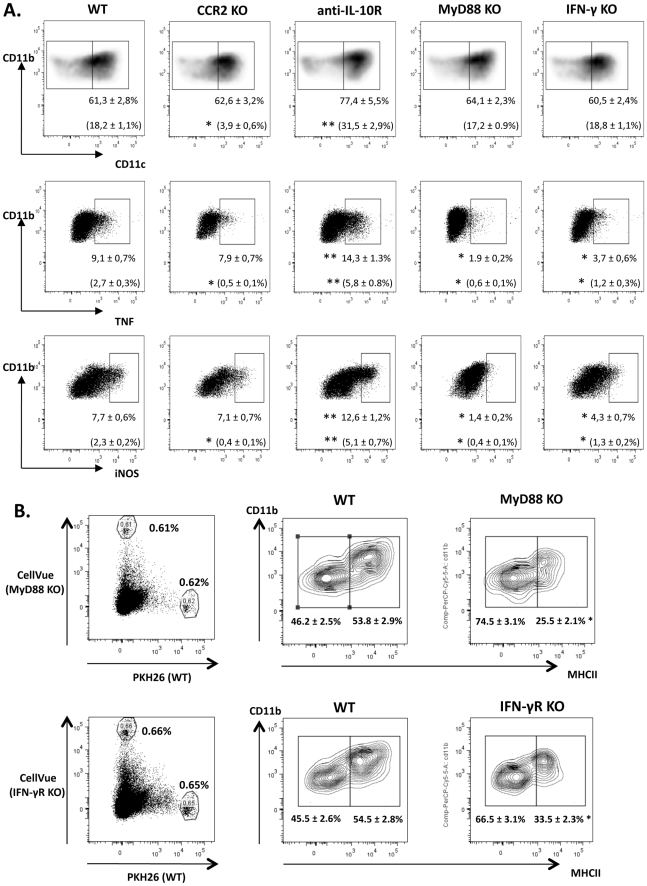
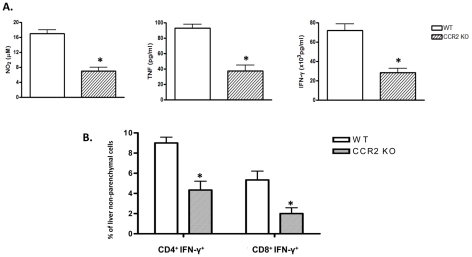
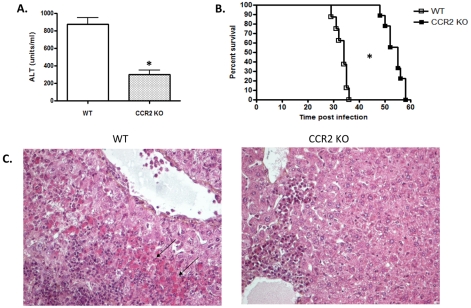
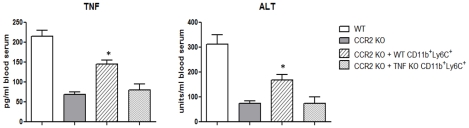
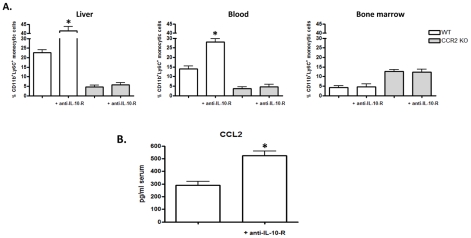
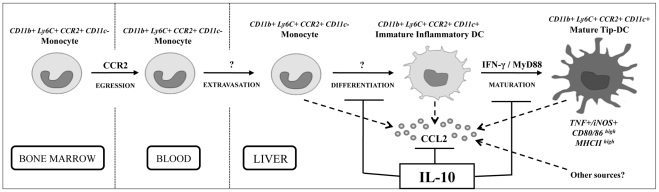
The development of classically activated monocytic cells (M1) is a prerequisite for effective elimination of parasites, including African trypanosomes. However, persistent activation of M1 that produce pathogenic molecules such as TNF and NO contributes to the development of trypanosome infection-associated tissue injury including liver cell necrosis in experimental mouse models. Aiming to identify mechanisms involved in regulation of M1 activity, we have recently documented that during Trypanosoma brucei infection, CD11b(+)Ly6C(+)CD11c(+) TNF and iNOS producing DCs (Tip-DCs) represent the major pathogenic M1 liver subpopulation. By using gene expression analyses, KO mice and cytokine neutralizing antibodies, we show here that the conversion of CD11b(+)Ly6C(+) monocytic cells to pathogenic Tip-DCs in the liver of T. brucei infected mice consists of a three-step process including (i) a CCR2-dependent but CCR5- and Mif-independent step crucial for emigration of CD11b(+)Ly6C(+) monocytic cells from the bone marrow but dispensable for their blood to liver migration; (ii) a differentiation step of liver CD11b(+)Ly6C(+) monocytic cells to immature inflammatory DCs (CD11c(+) but CD80/CD86/MHC-II(low)) which is IFN-gamma and MyD88 signaling independent; and (iii) a maturation step of inflammatory DCs to functional (CD80/CD86/MHC-II(high)) TNF and NO producing Tip-DCs which is IFN-gamma and MyD88 signaling dependent. Moreover, IL-10 could limit CCR2-mediated egression of CD11b(+)Ly6C(+) monocytic cells from the bone marrow by limiting Ccl2 expression by liver monocytic cells, as well as their differentiation and maturation to Tip-DCs in the liver, showing that IL-10 works at multiple levels to dampen Tip-DC mediated pathogenicity during T. brucei infection. A wide spectrum of liver diseases associates with alteration of monocyte recruitment, phenotype or function, which could be modulated by IL-10. Therefore, investigating the contribution of recruited monocytes to African trypanosome induced liver injury could potentially identify new targets to treat hepatic inflammation in general, and during parasite infection in particular.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
IL-27 Negatively Regulates Tip-DC Development during Infection.mBio. 2021 Feb 16;12(1):e03385-20. doi: 10.1128/mBio.03385-20. mBio. 2021. PMID: 33593983 Free PMC article.
-
IL-10 dampens TNF/inducible nitric oxide synthase-producing dendritic cell-mediated pathogenicity during parasitic infection.J Immunol. 2009 Jan 15;182(2):1107-18. doi: 10.4049/jimmunol.182.2.1107. J Immunol. 2009. PMID: 19124754
-
CD8 T cells regulate allergic contact dermatitis by modulating CCR2-dependent TNF/iNOS-expressing Ly6C+ CD11b+ monocytic cells.J Invest Dermatol. 2014 Mar;134(3):666-676. doi: 10.1038/jid.2013.403. Epub 2013 Sep 23. J Invest Dermatol. 2014. PMID: 24061165
-
Monocyte-mediated immune defense against murine Listeria monocytogenes infection.Adv Immunol. 2012;113:119-34. doi: 10.1016/B978-0-12-394590-7.00003-8. Adv Immunol. 2012. PMID: 22244581 Free PMC article. Review.
-
Estrogen receptors regulate an inflammatory pathway of dendritic cell differentiation: mechanisms and implications for immunity.Horm Behav. 2012 Aug;62(3):254-62. doi: 10.1016/j.yhbeh.2012.04.011. Epub 2012 Apr 25. Horm Behav. 2012. PMID: 22561458 Free PMC article. Review.
Cited by
-
Human and Mouse Mononuclear Phagocyte Networks: A Tale of Two Species?Front Immunol. 2015 Jun 25;6:330. doi: 10.3389/fimmu.2015.00330. eCollection 2015. Front Immunol. 2015. PMID: 26124761 Free PMC article. Review.
-
Interleukin expression after injury and the effects of interleukin-1 receptor antagonist.PLoS One. 2013 Aug 1;8(8):e71631. doi: 10.1371/journal.pone.0071631. Print 2013. PLoS One. 2013. PMID: 23936523 Free PMC article.
-
African Trypanosomiasis-Associated Anemia: The Contribution of the Interplay between Parasites and the Mononuclear Phagocyte System.Front Immunol. 2018 Feb 15;9:218. doi: 10.3389/fimmu.2018.00218. eCollection 2018. Front Immunol. 2018. PMID: 29497418 Free PMC article. Review.
-
Nitric oxide controls an inflammatory-like Ly6C(hi)PDCA1+ DC subset that regulates Th1 immune responses.J Leukoc Biol. 2011 Mar;89(3):443-55. doi: 10.1189/jlb.0610329. Epub 2010 Dec 22. J Leukoc Biol. 2011. PMID: 21178115 Free PMC article.
-
Interleukin-1 receptor antagonist modulates inflammation and scarring after ligament injury.Connect Tissue Res. 2014 Jun;55(3):177-86. doi: 10.3109/03008207.2014.906408. Epub 2014 Apr 18. Connect Tissue Res. 2014. PMID: 24649870 Free PMC article.
References
-
- Sternberg JM. Human African trypanosomiasis: clinical presentation and immune response. Parasite Immunol. 2004;26:469–476. - PubMed
-
- Naessens J. Bovine trypanotolerance: A natural ability to prevent severe anaemia and haemophagocytic syndrome? Int J Parasitol. 2006;36:521–528. - PubMed
-
- Bosschaerts T, Guilliams M, Stijlemans B, De Baetselier P, Beschin A. Understanding the role of monocytic cells in liver inflammation using parasite infection as a model. Immunobiology. 2009;214:737–747. - PubMed
-
- Drennan MB, Stijlemans B, Van den Abbeele J, Quesniaux VJ, Barkhuizen M, et al. The induction of a type 1 immune response following a Trypanosoma brucei infection is MyD88 dependent. J Immunol. 2005;175:2501–2509. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
