

Spatial cycles in G-protein crowd control
- PMID: 20717139
- PMCID: PMC2924655
- DOI: 10.1038/emboj.2010.184
Spatial cycles in G-protein crowd control
Abstract
The nature of living systems and their apparent resilience to the second law of thermodynamics has been the subject of extensive investigation and imaginative speculation. The segregation and compartmentalization of proteins is one manifestation of this departure from equilibrium conditions; the effect of which is now beginning to be elucidated. This should not come as a surprise, as even a cursory inspection of cellular processes reveals the large amount of energetic cost borne to maintain cell-scale patterns, separations and gradients of molecules. The G-proteins, kinases, calcium-responsive proteins have all been shown to contain reaction cycles that are inherently coupled to their signalling activities. G-proteins represent an important and diverse toolset used by cells to generate cellular asymmetries. Many small G-proteins in particular, are dynamically acylated to modify their membrane affinities, or localized in an activity-dependent manner, thus manipulating the mobility modes of these proteins beyond pure diffusion and leading to finely tuned steady state partitioning into cellular membranes. The rates of exchange of small G-proteins over various compartments, as well as their steady state distributions enrich and diversify the landscape of possibilities that GTPase-dependent signalling networks can display over cellular dimensions. The chemical manipulation of spatial cycles represents a new approach for the modulation of cellular signalling with potential therapeutic benefits.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
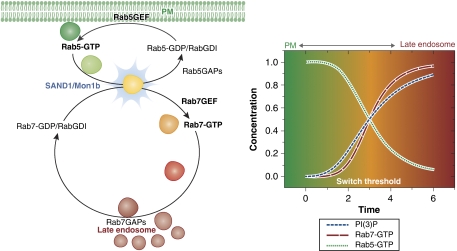
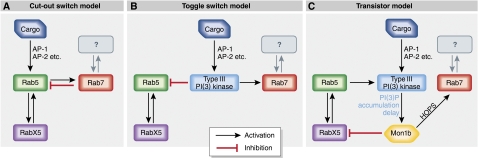
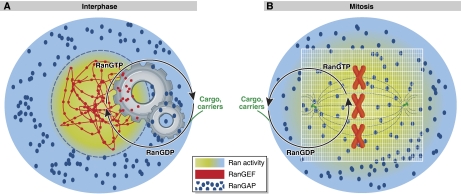
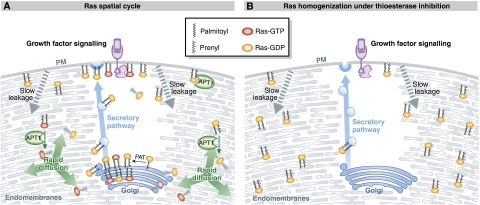
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
