

Review
doi: 10.1038/emboj.2010.182.
Spatial organization of adhesion: force-dependent regulation and function in tissue morphogenesis
Affiliations
- PMID: 20717145
- PMCID: PMC2924654
- DOI: 10.1038/emboj.2010.182
Item in Clipboard
Review
Spatial organization of adhesion: force-dependent regulation and function in tissue morphogenesis
EMBO J.
.
Abstract
Integrin- and cadherin-mediated adhesion is central for cell and tissue morphogenesis, allowing cells and tissues to change shape without loosing integrity. Studies predominantly in cell culture showed that mechanosensation through adhesion structures is achieved by force-mediated modulation of their molecular composition. The specific molecular composition of adhesion sites in turn determines their signalling activity and dynamic reorganization. Here, we will review how adhesion sites respond to mecanical stimuli, and how spatially and temporally regulated signalling from different adhesion sites controls cell migration and tissue morphogenesis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
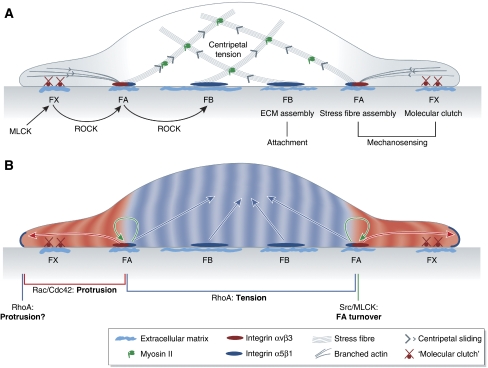
Force-dependent remodelling of integrin-ECM adhesion site composition and signalling activity. (A) Tension defines the molecular composition and size of integrin-ECM adhesion sites. Assembly of nascent integrin αvβ3-containing focal complexes (FXs) at the cell edge is driven by MLCK. FXs interact with sliding actin filaments through a ‘molecular clutch', enabling tension-dependent maturation into focal adhesions (FA), which is driven by ROCK. FAs trigger stress fibre assembly. ROCK-dependent maturation of integrin α5β1-containing fibrillar adhesions is necessary for ECM assembly and stable cell-substrate attachment. The maturation of integrin-ECM adhesion structures is dependent on tension created by myosin-driven centripetal sliding of actin cytoskeleton. (B) Molecular composition of integrin-ECM adhesion sites defines their signalling activities. RhoA is activated at the edge of peripheral cell protrusion by an unknown mechanism. Integrin αvβ3-containing FAs trigger peripheral Rac/Cdc42 activation that propagates distally and is necessary for stabilizing cell protrusions and downregulating peripheral RhoA activity. FAs also trigger activation of Src and MLCK that are essential for FA turnover during cell spreading and migration. Central integrin α5β1-containing adhesions reactivate RhoA, thereby contributing to the increase of centripetal actomyosin tension.
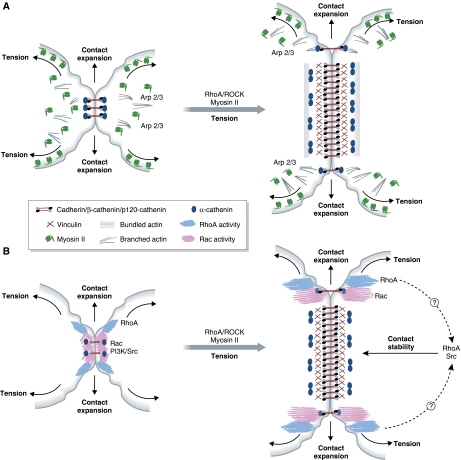
Force-dependent remodelling of cell–cell adhesion site composition and signalling activity. (A) Tension drives maturation of adherens junctions. Early adherens junctions are enriched in Arp 2/3 and branched actin. Actomyosin/ROCK-mediated tension is required for the expansion of adherens junction. As the junction expands, the molecular composition of its central part changes. The α-catenin dimerizes and dissociates from the cadherin–β-catenin complex and additional actin-binding adaptors, such as vinculin, are recruited. The α-catenin dimers are thought to modulate actin reorganization by promoting the assembly of junctional actin bundles. (B) Adherens junction maturation alters its signalling activity. Upon initial cadherin engagement at cell–cell contacts, Src/PI3 kinase as well as RhoA and Rac become activated. These signals propagate as the junction extends, and are downregulated at the mature, central region of the adherens junction. At the same time, RhoA and Src/PI3 kinase activities are essential for the stability of mature adherens junctions.
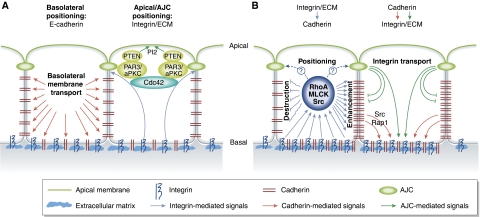
Establishment of epithelial cell polarity: crosstalk between cell–cell and cell–matrix adhesion. (A) Adhesion signals control epithelial polarization. Engagement of cadherins in a cell–cell contact triggers the transport of basolateral markers to the adherent membranes. Integrin-mediated adhesion of basal surface to the ECM is essential for apical localization of PTEN, PI2 and PAR complex and activation of Cdc42. Cdc42 activation in turn is responsible for the assembly of apical junction complexes (AJC) that separate the apical and basolateral domains. (B) Crosstalk between integrins and cadherins controls epithelial integrity. Integrin-mediated regulation of RhoA, Src and MLCK activities can lead to both an enhancement and destruction of adherens junctions. The specific effect depends on the ECM composition and the cell type. Cadherins also promote integrin-ECM adhesion through Src/Rap1 signalling and targeting of integrins to the basal membrane.
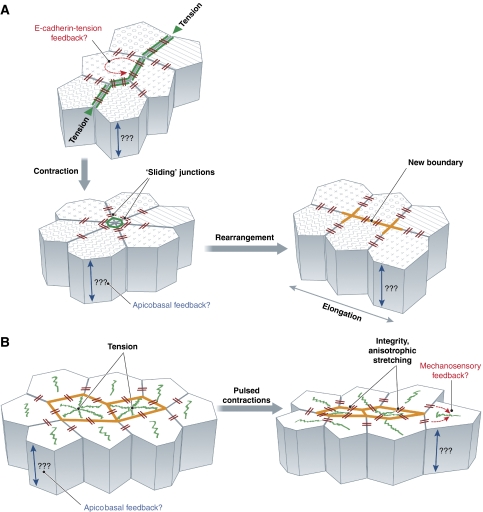
Remodelling of epithelial sheets in Drosophila embryos. (A) Planar cell rearrangement is dependent on actomyosin assembly at the apical junctions. Polarized assembly of junctional actomyosin filaments is driven by tension. Actomyosin-rich junction(s) contract, which in some cases can lead to a transient ‘rosette'-like cell assembly that subsequently dissolves through the polarized expansion of new boundaries. Polarized junction contraction and expansion drives germ band elongation. During planar cell rearrangements, E-cadherin accumulated in ‘spot junctions', which are connected to stable actin patches and linked to each other through a dynamic actin network ‘slide' along the boundaries. (B) Epithelial invagination is driven by pulsed contractions of a non-junctional actomyosin network. During epithelial invagination, coalescing actomyosin networks form at the apex of the invaginating cells. These networks are connected with E-cadherin-based spot junctions. Stochastic centripetal contractions of the apical actomyosin networks, followed by the stabilization of the contracted states, drives apical constriction. E-cadherin-based spot junctions are essential for maintaining tissue integrity and stabilizing anisotropic cell shape changes during the contraction pulses.
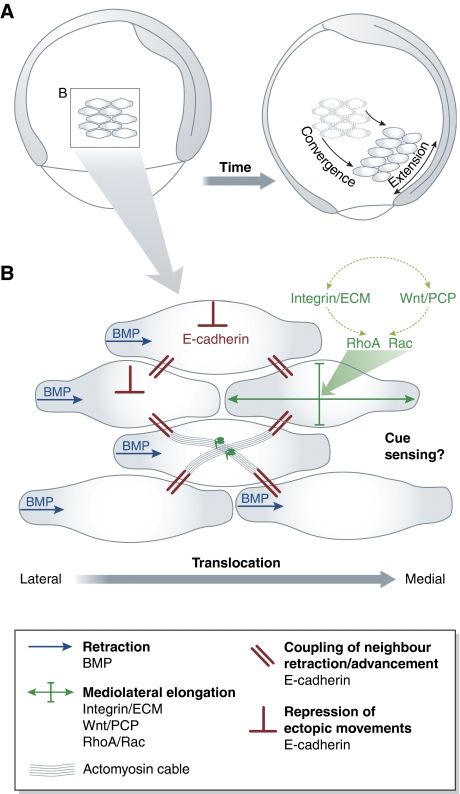
Collective migration of mesoderm progenitor cells in zebrafish gastrulation. (A) Convergence and extension movements of lateral mesoderm cells during zebrafish gastrulation. Lateral view of zebrafish embryo at onset and end of gastrulation. A group of mesoderm cells is highlighted. Cells migrate towards the dorsal side and undergo mediolateral cell intercalations. Both cell migration and intercalations contribute to convergence and extension of the forming body axis. (B) Collective migration of lateral mesoderm cells. Cell elongation is driven by integrin-ECM adhesion and Wnt/PCP signals, which together modulate Rho-family GTPase activity. BMP signalling polarizes protrusion formation in individual cells. E-cadherin-mediated cell cohesion mechanically couples and thereby aligns the movements of neighbouring cells. In addition, E-cadherin restricts movement of cells over each other.
References
-
- Anastasiadis PZ, Moon SY, Thoreson MA, Mariner DJ, Crawford HC, Zheng Y, Reynolds AB (2000) Inhibition of RhoA by p120 catenin. Nat Cell Biol 2: 637–644 - PubMed
-
- Arboleda-Estudillo Y, Krieg M, Stuhmer J, Licata NA, Muller DJ, Heisenberg CP (2010) Movement directionality in collective migration of germ layer progenitors. Curr Biol 20: 161–169 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
