

Proteomic characterization of human plasma high density lipoprotein fractionated by gel filtration chromatography
- PMID: 20718489
- PMCID: PMC3100175
- DOI: 10.1021/pr100520x
Proteomic characterization of human plasma high density lipoprotein fractionated by gel filtration chromatography
Abstract
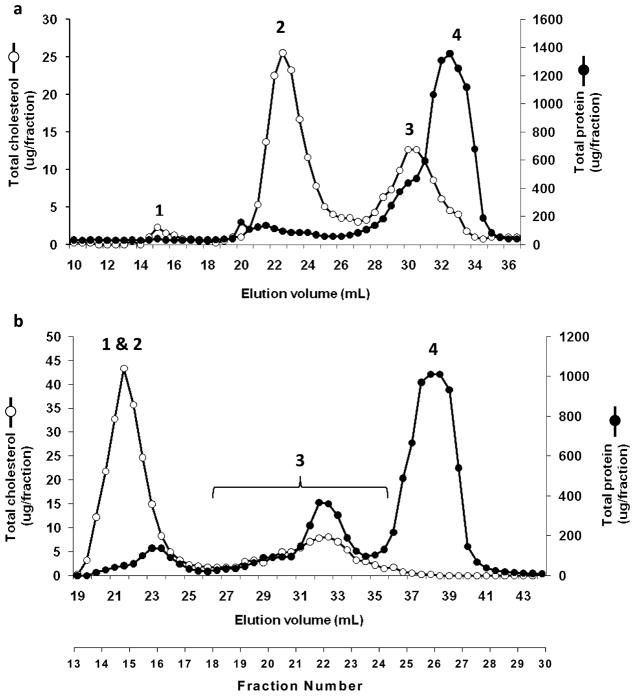
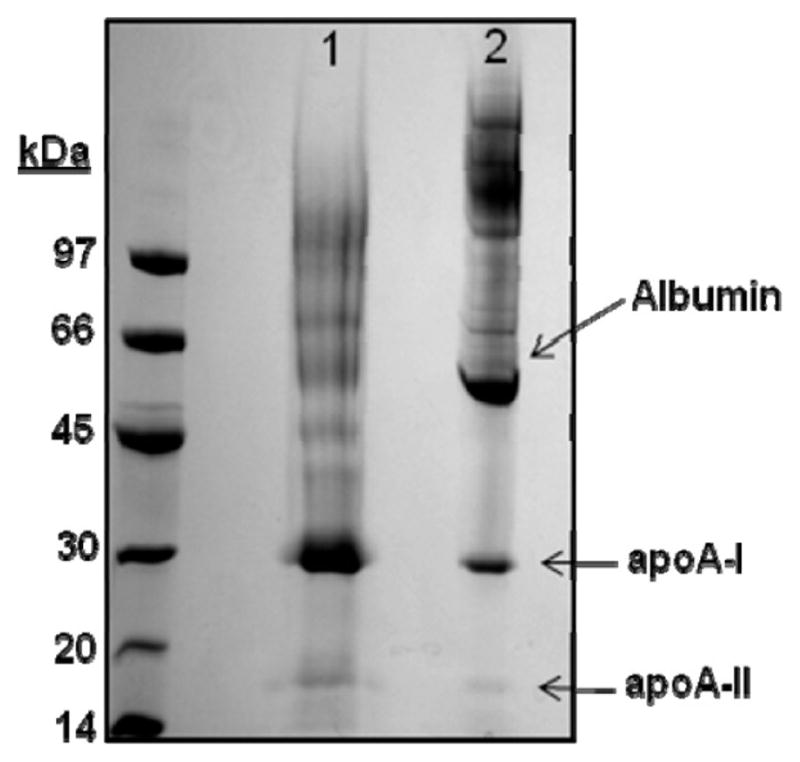
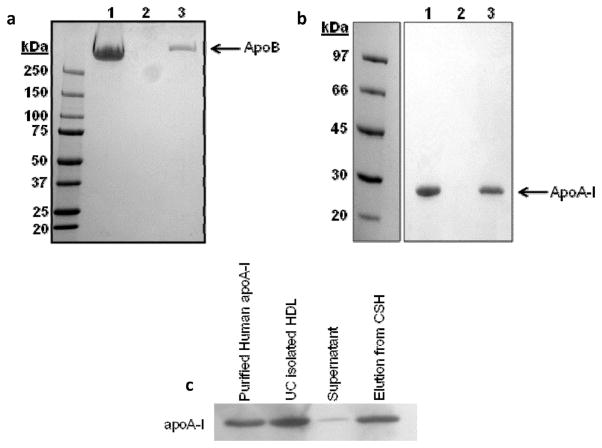
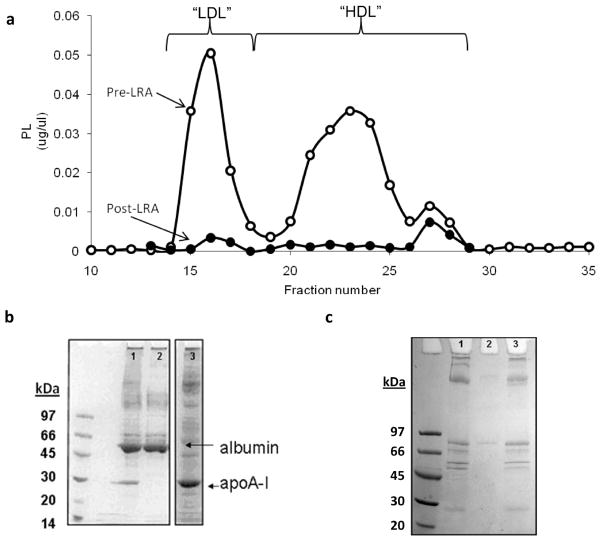
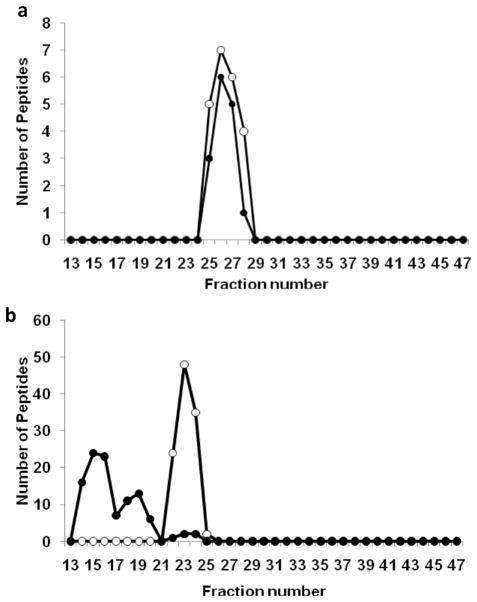
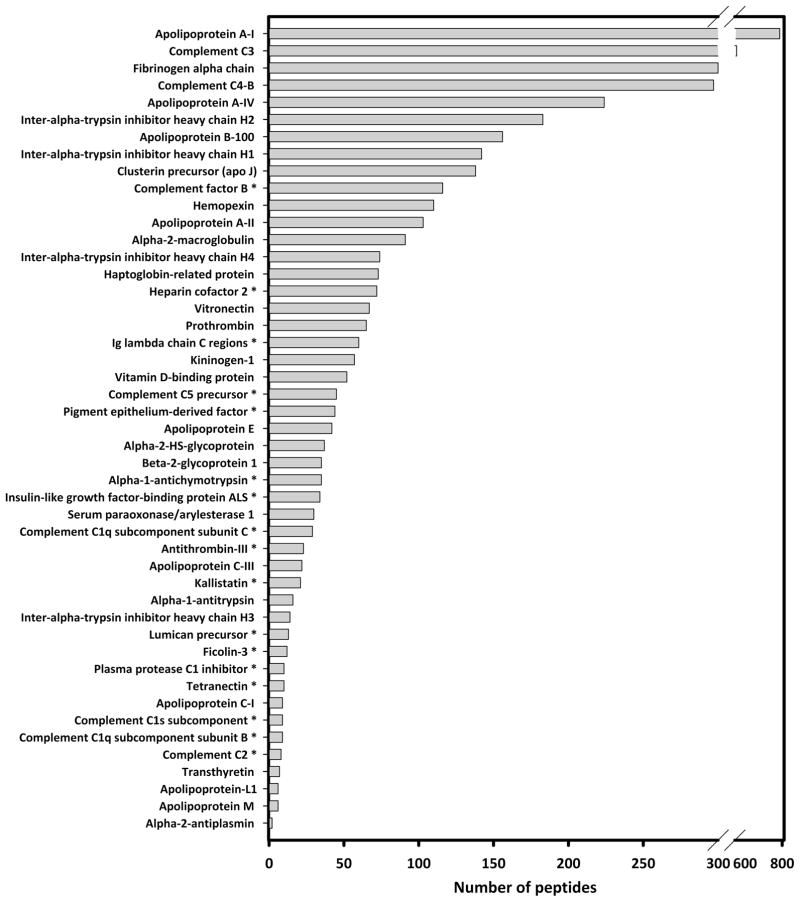
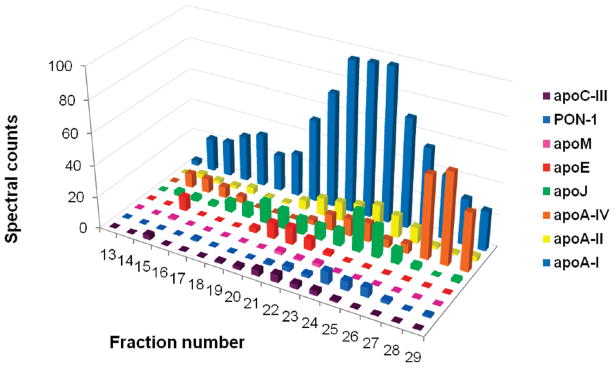
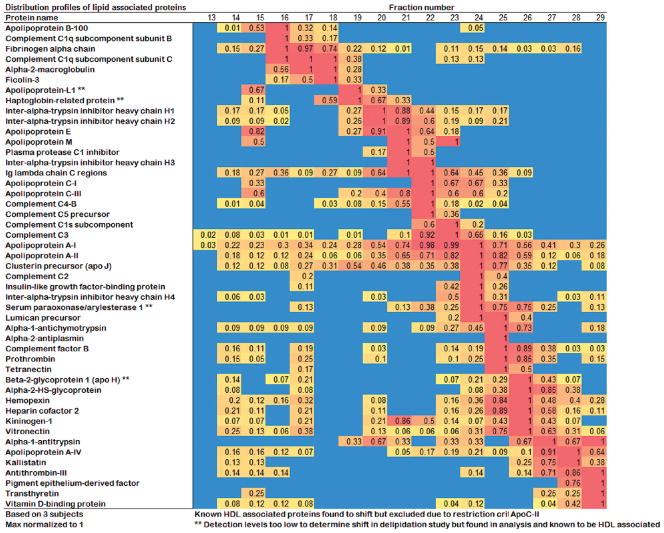
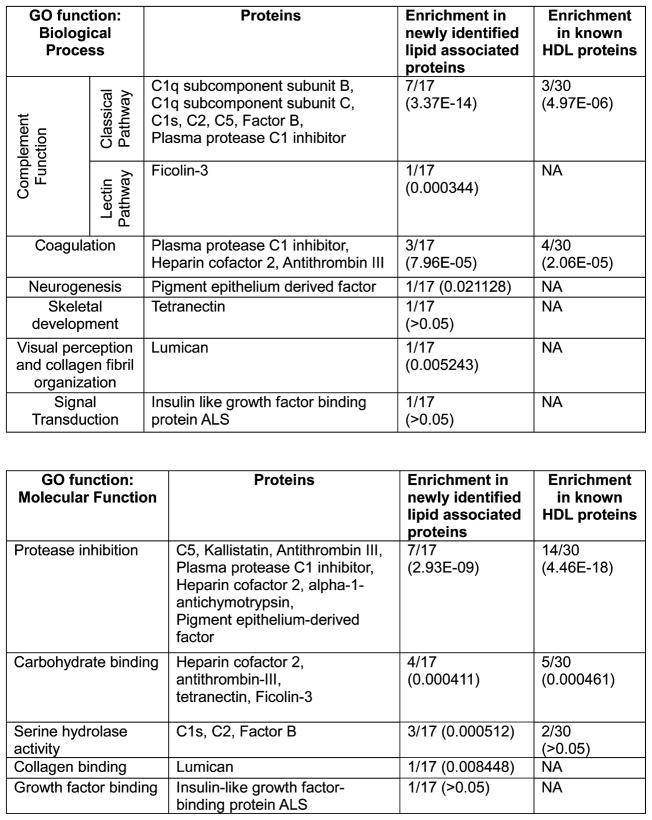
Plasma levels of high density lipoprotein cholesterol (HDL-C) are inversely proportional to the incidence of cardiovascular disease. Recent applications of modern proteomic technologies have identified upward of 50 distinct proteins associated with HDL particles with many of these newly discovered proteins implicating HDL in nonlipid transport processes including complement activation, acute phase response and innate immunity. However, almost all MS-based proteomic studies on HDL to date have utilized density gradient ultracentrifugation techniques for HDL isolation prior to analysis. These involve high shear forces and salt concentrations that can disrupt HDL protein interactions and alter particle function. Here, we used high-resolution size exclusion chromatography to fractionate normal human plasma to 17 phospholipid-containing subfractions. Then, using a phospholipid binding resin, we identified proteins that associate with lipoproteins of various sizes by electrospray ionization mass spectrometry. We identified 14 new phospholipid-associated proteins that migrate with traditionally defined HDL, several of which further support roles for HDL in complement regulation and protease inhibition. The increased fractionation inherent to this method allowed us to visualize HDL protein distribution across particle size with unprecedented resolution. The observed heterogeneity across subfractions suggests the presence of HDL particle subpopulations each with distinct protein components that may prove to impart distinct physiological functions.
Figures
References
-
- Scanu AM, Wisdom C. Serum lipoproteins structure and function. Annu Rev Biochem. 1972;41:703–730. - PubMed
-
- Franceschini G, Maderna P, Sirtori CR. Reverse cholesterol transport: physiology and pharmacology. Atherosclerosis. 1991;88(2–3):99–107. - PubMed
-
- Gordon T, Castelli WP, Hjortland MC, Kannel WB, Dawber TR. High density lipoprotein as a protective factor against coronary heart disease. Am J Med. 1977;62(5):707–714. - PubMed
-
- Gordon DJ, Probstfield JL, Garrison RJ, Neaton JD, Castelli WP, Knoke JD, Jacobs DR, Jr, Bangdiwala S, Tyroler HA. High-density lipoprotein cholesterol and cardiovascular disease. Four prospective American studies. Circulation. 1989;79(1):8–15. - PubMed
-
- Mackness MI, Arrol S, Durrington PN. Paraoxonase prevents accumulation of lipoperoxides in low-density lipoprotein. FEBS Lett. 1991;286(1–2):152–154. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
