

Human CD133-derived bone marrow stromal cells establish ectopic hematopoietic microenvironments in immunodeficient mice
- PMID: 20719235
- PMCID: PMC2942979
- DOI: 10.1016/j.bbrc.2010.08.040
Human CD133-derived bone marrow stromal cells establish ectopic hematopoietic microenvironments in immunodeficient mice
Abstract
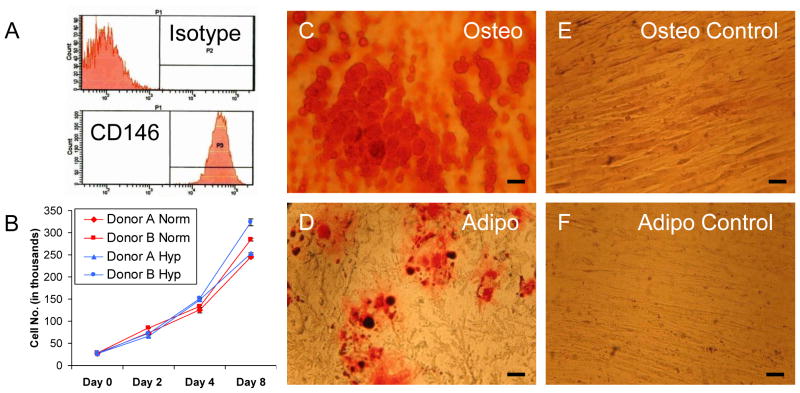
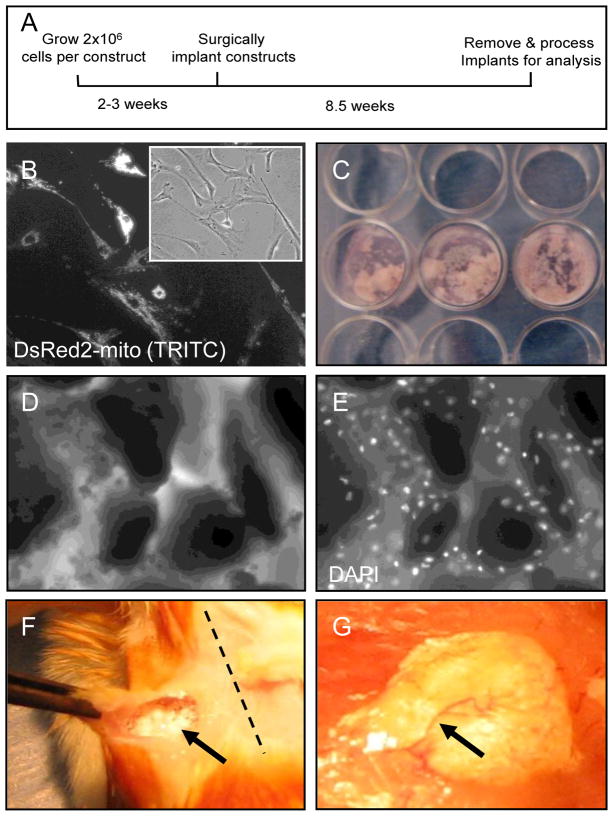
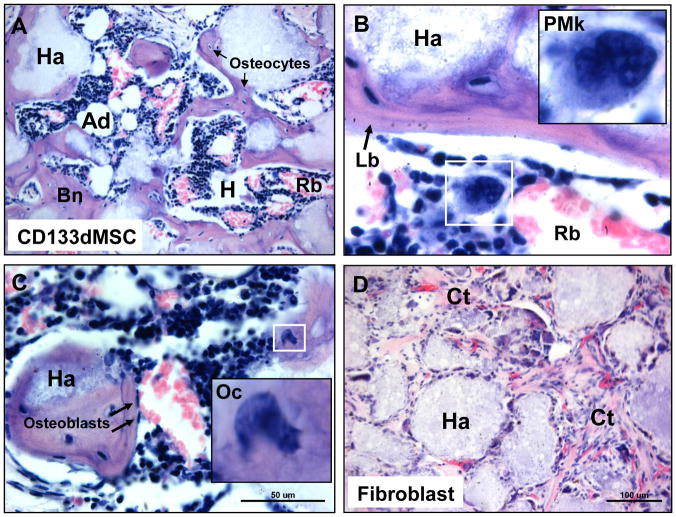
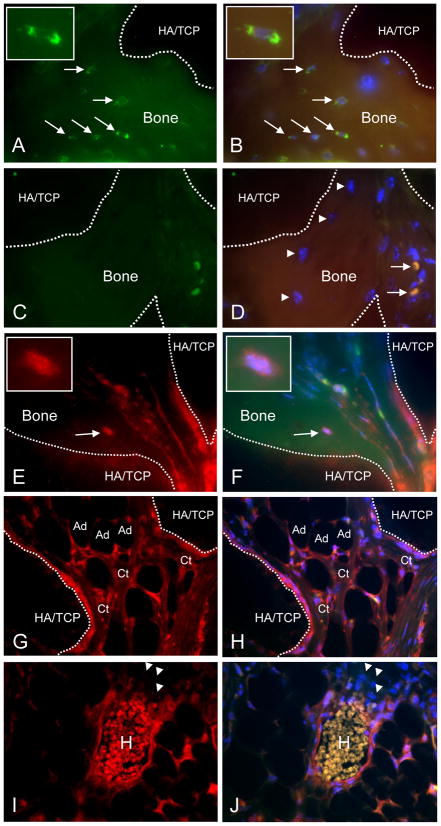
Cultured adherent bone marrow stromal cells (BMSCs) are capable of forming ectopic hematopoietic microenvironments (HMEs) in immunodeficient mice. However, the cell surface phenotype of the native bone marrow stem/progenitor cell that gives rise to BMSCs that support hematopoiesis remains poorly defined. We recently reported the derivation of human BMSC-like cells (CD133BMSCs) by magnetic cell sorting against Prominin-1 (CD133), an epitope expressed by embryonic, fetal, and adult stem cells. Here we demonstrate that CD133BMSCs are capable of forming ectopic HMEs. Cultured adherent CD133BMSCs derived from sorted CD133-positive cells lacked CD133 expression, but were uniformly positive for CD146, an epitope recently described to identify self-renewing osteoprogenitor cells that could transfer the HME. CD133BMSCs were genetically-tagged by lentivirus, expanded, and seeded into HA/TCP/fibrin constructs that were implanted subcutaneously. After 60days, CD133BMSCs produced human osteocytes, osteoblasts, adipocytes, and reticular cells that supported murine hematopoiesis. CD133BMSCs that were not transduced with lentivirus also formed HMEs. Control constructs seeded with human dermal fibroblasts formed connective tissue, but failed to form HMEs. Our data indicate that CD133 expression identifies a native human bone marrow stem/progenitor cell that gives rise to BMSCs capable of forming the HME.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Wilson A, Trumpp A. Bone-marrow haematopoietic-stem-cell niches. Nat Rev Immunol. 2006;6:93–106. - PubMed
-
- Zhang J, Niu C, Ye L, et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature. 2003;425:836–41. - PubMed
-
- Kiel MJ, Yilmaz OH, Iwashita T, et al. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell. 2005;121:1109–21. - PubMed
-
- Calvi LM, Adams GB, Weibrecht KW, et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature. 2003;425:841–6. - PubMed
-
- Scadden DT. The stem-cell niche as an entity of action. Nature. 2006;441:1075–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
