

Sprouty proteins inhibit receptor-mediated activation of phosphatidylinositol-specific phospholipase C
- PMID: 20719962
- PMCID: PMC2947483
- DOI: 10.1091/mbc.E10-02-0123
Sprouty proteins inhibit receptor-mediated activation of phosphatidylinositol-specific phospholipase C
Abstract
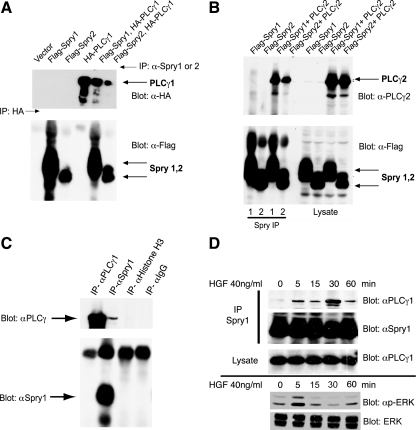
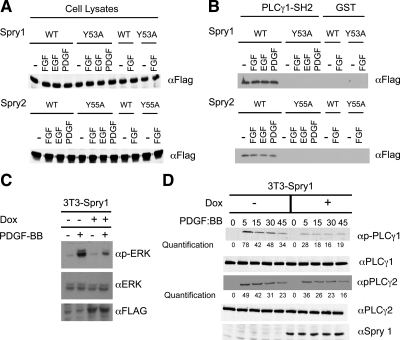
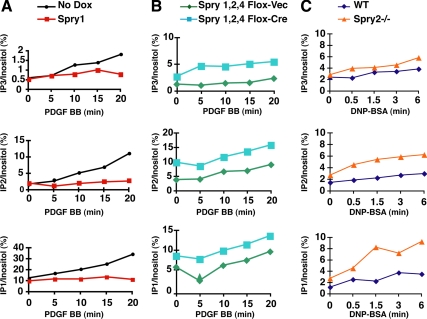
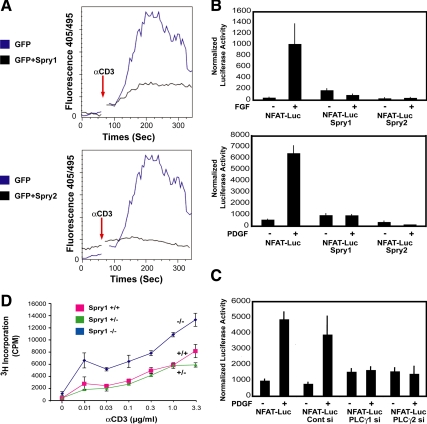
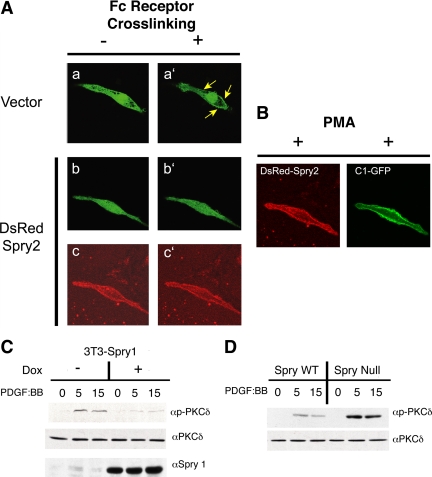
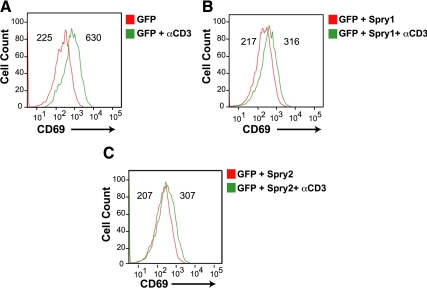
Sprouty (Spry) proteins are negative regulators of receptor tyrosine kinase signaling; however, their exact mechanism of action remains incompletely understood. We identified phosphatidylinositol-specific phospholipase C (PLC)-γ as a partner of the Spry1 and Spry2 proteins. Spry-PLCγ interaction was dependent on the Src homology 2 domain of PLCγ and a conserved N-terminal tyrosine residue in Spry1 and Spry2. Overexpression of Spry1 and Spry2 was associated with decreased PLCγ phosphorylation and decreased PLCγ activity as measured by production of inositol (1,4,5)-triphosphate (IP(3)) and diacylglycerol, whereas cells deficient for Spry1 or Spry1, -2, and -4 showed increased production of IP(3) at baseline and further increased in response to growth factor signals. Overexpression of Spry 1 or Spry2 or small-interfering RNA-mediated knockdown of PLCγ1 or PLCγ2 abrogated the activity of a calcium-dependent reporter gene, suggesting that Spry inhibited calcium-mediated signaling downstream of PLCγ. Furthermore, Spry overexpression in T-cells, which are highly dependent on PLCγ activity and calcium signaling, blocked T-cell receptor-mediated calcium release. Accordingly, cultured T-cells from Spry1 gene knockout mice showed increased proliferation in response to T-cell receptor stimulation. These data highlight an important action of Spry, which may allow these proteins to influence signaling through multiple receptors.
Figures
References
-
- Abe M., Naski M. C. Regulation of sprouty expression by PLCγ and calcium-dependent signals. Biochem. Biophys. Res. Commun. 2004;323:1040–1047. - PubMed
-
- Ayada T., Taniguchi K., Okamoto F., Kato R., Komune S., Takaesu G., Yoshimura A. Sprouty4 negatively regulates protein kinase C activation by inhibiting phosphatidylinositol 4,5-biphosphate hydrolysis. Oncogene. 2009;28:1076–1088. - PubMed
-
- Basson M. A., et al. Sprouty1 is a critical regulator of GDNF/RET-mediated kidney induction. Dev. Cell. 2005;8:229–239. - PubMed
-
- Basson M. A., Watson-Johnson J., Shakya R., Akbulut S., Hyink D., Costantini F. D., Wilson P. D., Mason I. J., Licht J. D. Branching morphogenesis of the ureteric epithelium during kidney development is coordinated by the opposing functions of GDNF and Sprouty1. Dev. Biol. 2006;299:466–477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
