

Structure and function of the hetero-oligomeric cysteine synthase complex in plants
- PMID: 20720017
- PMCID: PMC2963375
- DOI: 10.1074/jbc.M110.157446
Structure and function of the hetero-oligomeric cysteine synthase complex in plants
Abstract
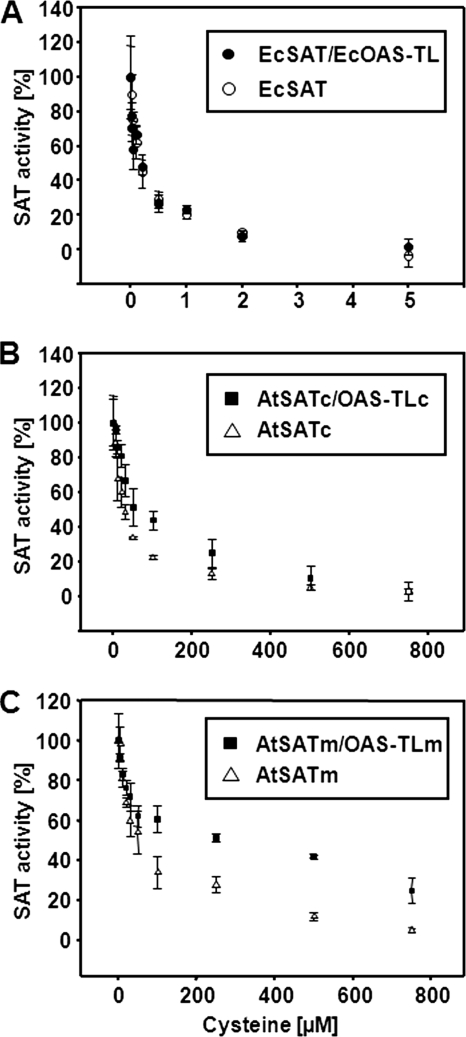
Cysteine synthesis in bacteria and plants is catalyzed by serine acetyltransferase (SAT) and O-acetylserine (thiol)-lyase (OAS-TL), which form the hetero-oligomeric cysteine synthase complex (CSC). In plants, but not in bacteria, the CSC is assumed to control cellular sulfur homeostasis by reversible association of the subunits. Application of size exclusion chromatography, analytical ultracentrifugation, and isothermal titration calorimetry revealed a hexameric structure of mitochondrial SAT from Arabidopsis thaliana (AtSATm) and a 2:1 ratio of the OAS-TL dimer to the SAT hexamer in the CSC. Comparable results were obtained for the composition of the cytosolic SAT from A. thaliana (AtSATc) and the cytosolic SAT from Glycine max (Glyma16g03080, GmSATc) and their corresponding CSCs. The hexameric SAT structure is also supported by the calculated binding energies between SAT trimers. The interaction sites of dimers of AtSATm trimers are identified using peptide arrays. A negative Gibbs free energy (ΔG = -33 kcal mol(-1)) explains the spontaneous formation of the AtCSCs, whereas the measured SAT:OAS-TL affinity (K(D) = 30 nm) is 10 times weaker than that of bacterial CSCs. Free SAT from bacteria is >100-fold more sensitive to feedback inhibition by cysteine than AtSATm/c. The sensitivity of plant SATs to cysteine is further decreased by CSC formation, whereas the feedback inhibition of bacterial SAT by cysteine is not affected by CSC formation. The data demonstrate highly similar quaternary structures of the CSCs from bacteria and plants but emphasize differences with respect to the affinity of CSC formation (K(D)) and the regulation of cysteine sensitivity of SAT within the CSC.
Figures





Similar articles
-
Functional analysis of the cysteine synthase protein complex from plants: structural, biochemical and regulatory properties.J Plant Physiol. 2006 Feb;163(3):273-86. doi: 10.1016/j.jplph.2005.11.013. Epub 2005 Dec 28. J Plant Physiol. 2006. PMID: 16386330 Review.
-
Mitochondrial cysteine synthase complex regulates O-acetylserine biosynthesis in plants.J Biol Chem. 2012 Aug 10;287(33):27941-7. doi: 10.1074/jbc.M112.372656. Epub 2012 Jun 22. J Biol Chem. 2012. PMID: 22730323 Free PMC article.
-
Allosterically gated enzyme dynamics in the cysteine synthase complex regulate cysteine biosynthesis in Arabidopsis thaliana.Structure. 2012 Feb 8;20(2):292-302. doi: 10.1016/j.str.2011.11.019. Structure. 2012. PMID: 22325778
-
Dominant-negative modification reveals the regulatory function of the multimeric cysteine synthase protein complex in transgenic tobacco.Plant Cell. 2007 Feb;19(2):625-39. doi: 10.1105/tpc.106.043125. Epub 2007 Feb 9. Plant Cell. 2007. PMID: 17293569 Free PMC article.
-
Molecular and biochemical analysis of serine acetyltransferase and cysteine synthase towards sulfur metabolic engineering in plants.Amino Acids. 2002;22(3):231-43. doi: 10.1007/s007260200011. Amino Acids. 2002. PMID: 12083067 Review.
Cited by
-
Modulation of Escherichia coli serine acetyltransferase catalytic activity in the cysteine synthase complex.FEBS Lett. 2017 May;591(9):1212-1224. doi: 10.1002/1873-3468.12630. Epub 2017 Apr 17. FEBS Lett. 2017. PMID: 28337759 Free PMC article.
-
Drought stress in maize causes differential acclimation responses of glutathione and sulfur metabolism in leaves and roots.BMC Plant Biol. 2016 Nov 9;16(1):247. doi: 10.1186/s12870-016-0940-z. BMC Plant Biol. 2016. PMID: 27829370 Free PMC article.
-
Metabolism and Regulatory Functions of O-Acetylserine, S-Adenosylmethionine, Homocysteine, and Serine in Plant Development and Environmental Responses.Front Plant Sci. 2021 May 7;12:643403. doi: 10.3389/fpls.2021.643403. eCollection 2021. Front Plant Sci. 2021. PMID: 34025692 Free PMC article. Review.
-
The Interplay between Hydrogen Sulfide and Phytohormone Signaling Pathways under Challenging Environments.Int J Mol Sci. 2022 Apr 12;23(8):4272. doi: 10.3390/ijms23084272. Int J Mol Sci. 2022. PMID: 35457090 Free PMC article. Review.
-
The response of Caenorhabditis elegans to hydrogen sulfide and hydrogen cyanide.Genetics. 2011 Oct;189(2):521-32. doi: 10.1534/genetics.111.129841. Epub 2011 Aug 11. Genetics. 2011. PMID: 21840852 Free PMC article.
References
-
- Kredich N. M., Becker M. A., Tomkins G. M. (1969) J. Biol. Chem. 244, 2428–2439 - PubMed
-
- Wirtz M., Hell R. (2006) J. Plant Physiol. 163, 273–286 - PubMed
-
- Droux M., Ruffet M. L., Douce R., Job D. (1998) Eur. J. Biochem. 255, 235–245 - PubMed
-
- Kredich N. M. (1996) in Escherichia coli and Salmonella Typhimurium. Cellular and Molecular Biology (Neidhardt F. C., Curtiss R., Ingraham J. L., Lin E. C., Low K. B., Magasanik B., Reznikoff W. S., Riley M., Schaechter M., Umberger E. eds) pp. 514–527, American Society for Microbiology, Washington, D. C
-
- Sirko A., Błaszczyk A., Liszewska F. (2004) J. Exp. Bot. 55, 1881–1888 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
