

Decreased rhythmic GABAergic septal activity and memory-associated theta oscillations after hippocampal amyloid-beta pathology in the rat
- PMID: 20720106
- PMCID: PMC6633464
- DOI: 10.1523/JNEUROSCI.6284-09.2010
Decreased rhythmic GABAergic septal activity and memory-associated theta oscillations after hippocampal amyloid-beta pathology in the rat
Abstract
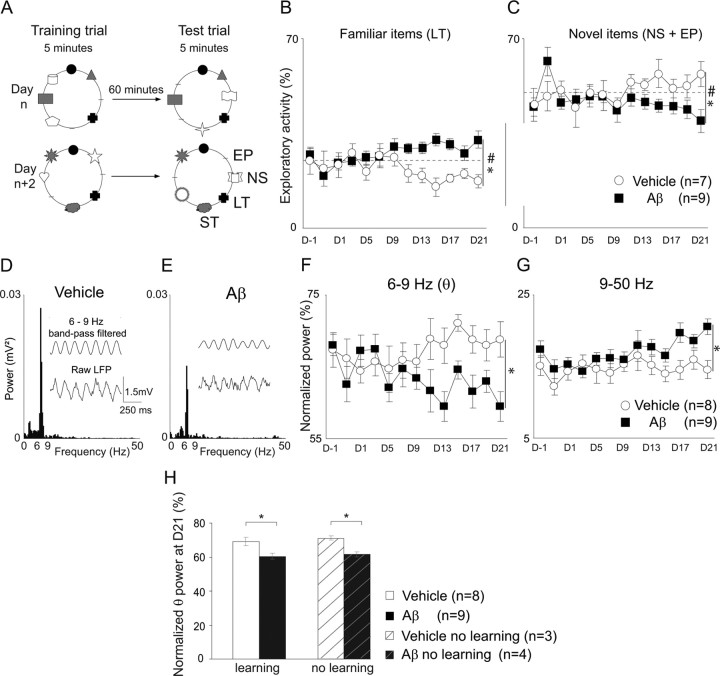
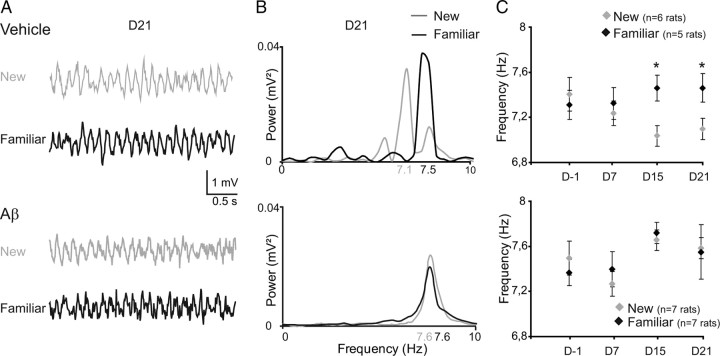
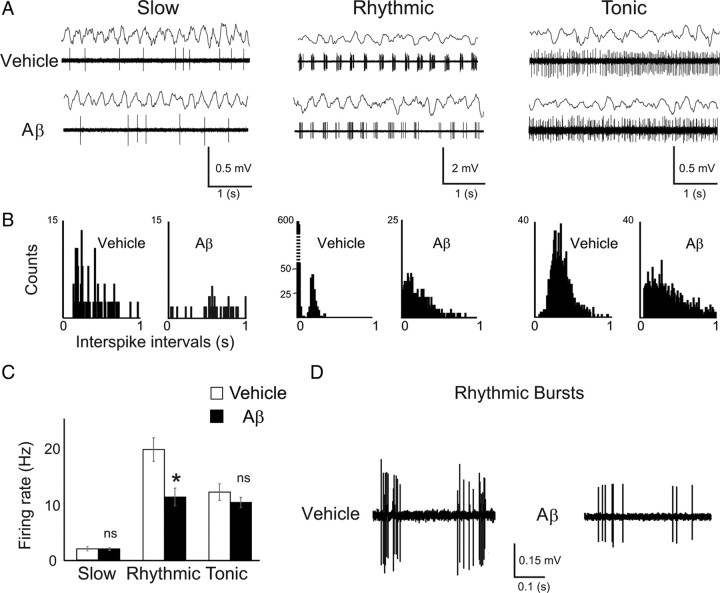
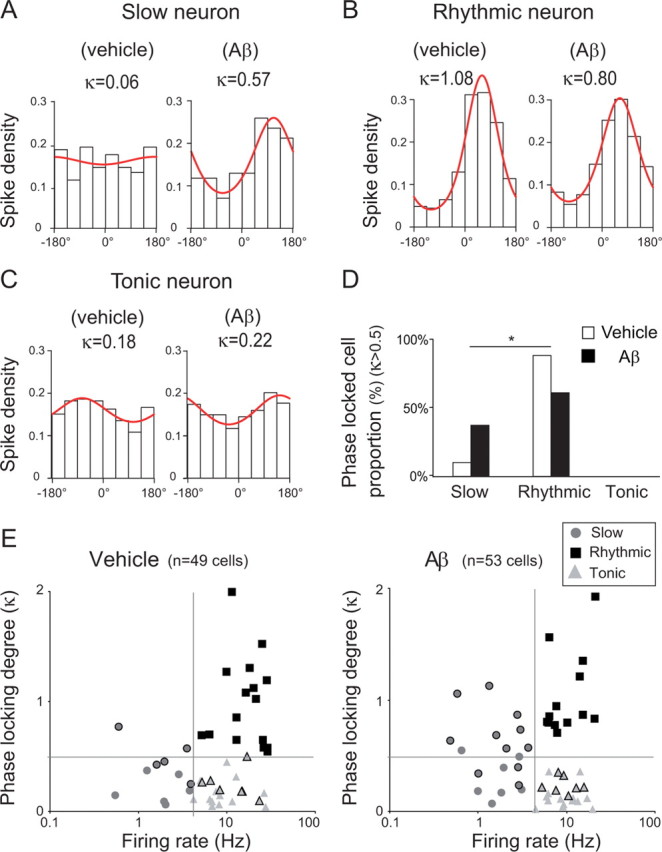
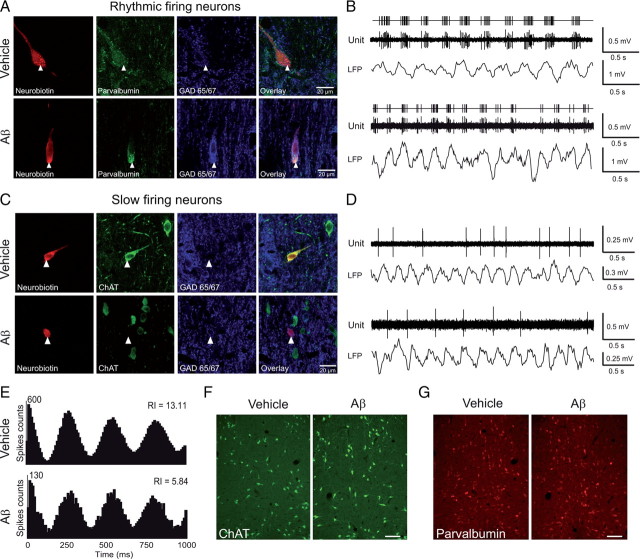
The memory deficits associated with Alzheimer's disease result to a great extent from hippocampal network dysfunction. The coordination of this network relies on theta (symbol) oscillations generated in the medial septum. Here, we investigated in rats the impact of hippocampal amyloid beta (Abeta) injections on the physiological and cognitive functions that depend on the septohippocampal system. Hippocampal Abeta injections progressively impaired behavioral performances, the associated hippocampal theta power, and theta frequency response in a visuospatial recognition test. These alterations were associated with a specific reduction in the firing of the identified rhythmic bursting GABAergic neurons responsible for the propagation of the theta rhythm to the hippocampus, but without loss of medial septal neurons. Such results indicate that hippocampal Abeta treatment leads to a specific functional depression of inhibitory projection neurons of the medial septum, resulting in the functional impairment of the temporal network.
Conflict of interest statement
The authors declare that they have no actual or potential conflict of interest, financial or otherwise, related to the present work. The data contained in this work were not submitted elsewhere. Experimental procedures and the manuscript were approved by all authors.
Figures
Similar articles
-
Medial septal beta-amyloid 1-40 injections alter septo-hippocampal anatomy and function.Neurobiol Aging. 2010 Jan;31(1):46-57. doi: 10.1016/j.neurobiolaging.2008.05.006. Epub 2008 Jun 10. Neurobiol Aging. 2010. PMID: 18547680 Free PMC article.
-
GABAergic Medial Septal Neurons with Low-Rhythmic Firing Innervating the Dentate Gyrus and Hippocampal Area CA3.J Neurosci. 2019 Jun 5;39(23):4527-4549. doi: 10.1523/JNEUROSCI.3024-18.2019. Epub 2019 Mar 29. J Neurosci. 2019. PMID: 30926750 Free PMC article.
-
Spatio-temporal specialization of GABAergic septo-hippocampal neurons for rhythmic network activity.Brain Struct Funct. 2018 Jun;223(5):2409-2432. doi: 10.1007/s00429-018-1626-0. Epub 2018 Mar 3. Brain Struct Funct. 2018. PMID: 29500537 Free PMC article.
-
Disturbances of septohippocampal theta oscillations in the epileptic brain: reasons and consequences.Exp Neurol. 2013 Sep;247:314-27. doi: 10.1016/j.expneurol.2013.01.029. Epub 2013 Feb 4. Exp Neurol. 2013. PMID: 23384663 Review.
-
Brainstem-diencephalo-septohippocampal systems controlling the theta rhythm of the hippocampus.Neuroscience. 1997 Dec;81(4):893-926. doi: 10.1016/s0306-4522(97)00239-x. Neuroscience. 1997. PMID: 9330355 Review.
Cited by
-
Effects of the γ-secretase inhibitor semagacestat on hippocampal neuronal network oscillation.Front Pharmacol. 2013 Jun 14;4:72. doi: 10.3389/fphar.2013.00072. eCollection 2013. Front Pharmacol. 2013. PMID: 23785331 Free PMC article.
-
Amyloid beta inhibits olfactory bulb activity and the ability to smell.PLoS One. 2013 Sep 26;8(9):e75745. doi: 10.1371/journal.pone.0075745. eCollection 2013. PLoS One. 2013. PMID: 24086624 Free PMC article.
-
Enhanced accumulation of N-terminally truncated Aβ with and without pyroglutamate-11 modification in parvalbumin-expressing GABAergic neurons in idiopathic and dup15q11.2-q13 autism.Acta Neuropathol Commun. 2020 Apr 28;8(1):58. doi: 10.1186/s40478-020-00923-8. Acta Neuropathol Commun. 2020. PMID: 32345355 Free PMC article.
-
Integrative EEG biomarkers predict progression to Alzheimer's disease at the MCI stage.Front Aging Neurosci. 2013 Oct 3;5:58. doi: 10.3389/fnagi.2013.00058. eCollection 2013. Front Aging Neurosci. 2013. PMID: 24106478 Free PMC article.
-
Activation of G-protein-gated inwardly rectifying potassium (Kir3/GirK) channels rescues hippocampal functions in a mouse model of early amyloid-β pathology.Sci Rep. 2017 Nov 7;7(1):14658. doi: 10.1038/s41598-017-15306-8. Sci Rep. 2017. PMID: 29116174 Free PMC article.
References
-
- Akiyama H, Barger S, Barnum S, Bradt B, Bauer J, Cole GM, Cooper NR, Eikelenboom P, Emmerling M, Fiebich BL, Finch CE, Frautschy S, Griffin WS, Hampel H, Hull M, Landreth G, Lue L, Mrak R, Mackenzie IR, McGeer PL, et al. Inflammation and Alzheimer's disease. Neurobiol Aging. 2000;21:383–421. - PMC - PubMed
-
- Ambrée O, Touma C, Görtz N, Keyvani K, Paulus W, Palme R, Sachser N. Activity changes and marked stereotypic behavior precede Abeta pathology in TgCRND8 Alzheimer mice. Neurobiol Aging. 2006;27:955–964. - PubMed
-
- Apartis E, Poindessous-Jazat FR, Lamour YA, Bassant MH. Loss of rhythmically bursting neurons in rat medial septum following selective lesion of septohippocampal cholinergic system. J Neurophysiol. 1998;79:1633–1642. - PubMed
-
- Arkhipov V, Kulesskaja N, Lebedev D. Behavioral perseveration and impairment of long-term memory in rats after intrahippocampal injection of kainic acid in subconvulsive dose. Pharmacol Biochem Behav. 2008;88:299–305. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical