

Spectral integration in primary auditory cortex attributable to temporally precise convergence of thalamocortical and intracortical input
- PMID: 20720119
- PMCID: PMC6633479
- DOI: 10.1523/JNEUROSCI.0689-10.2010
Spectral integration in primary auditory cortex attributable to temporally precise convergence of thalamocortical and intracortical input
Abstract
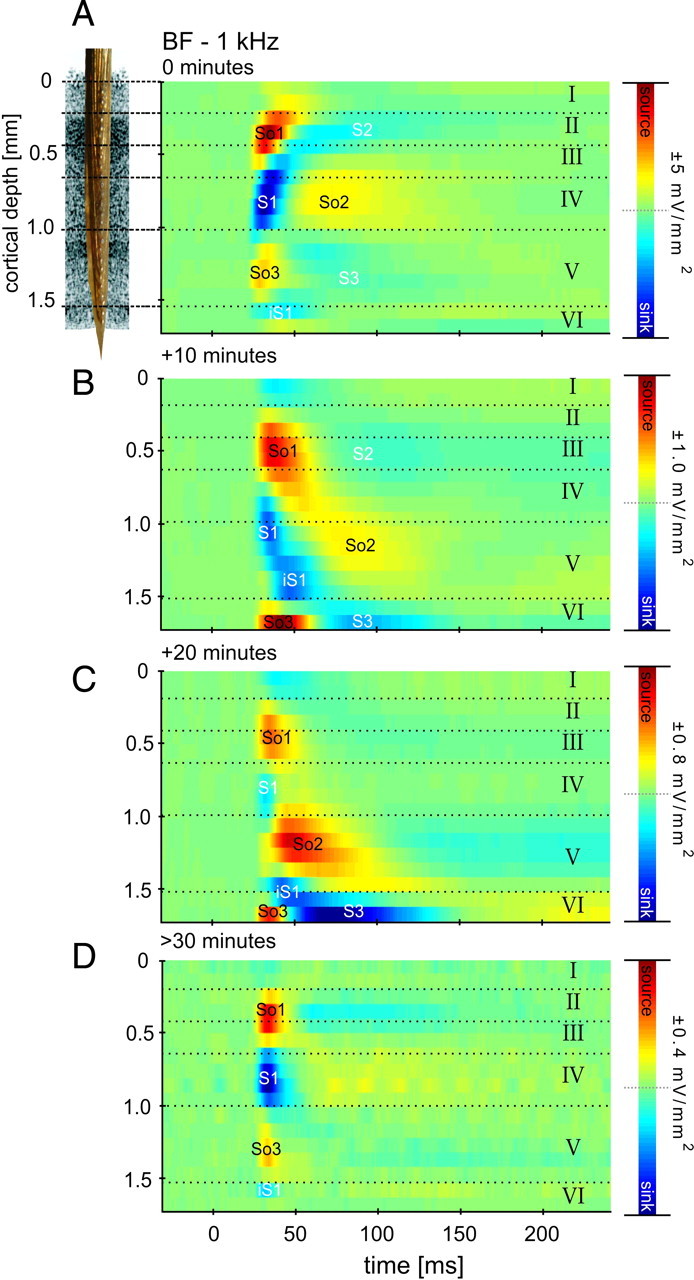
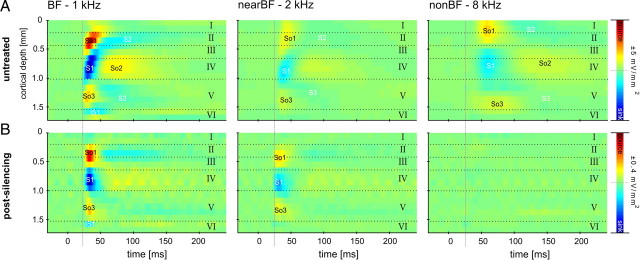
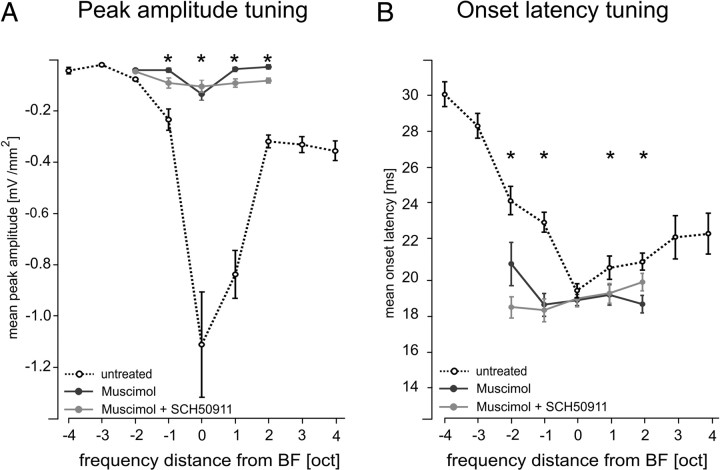
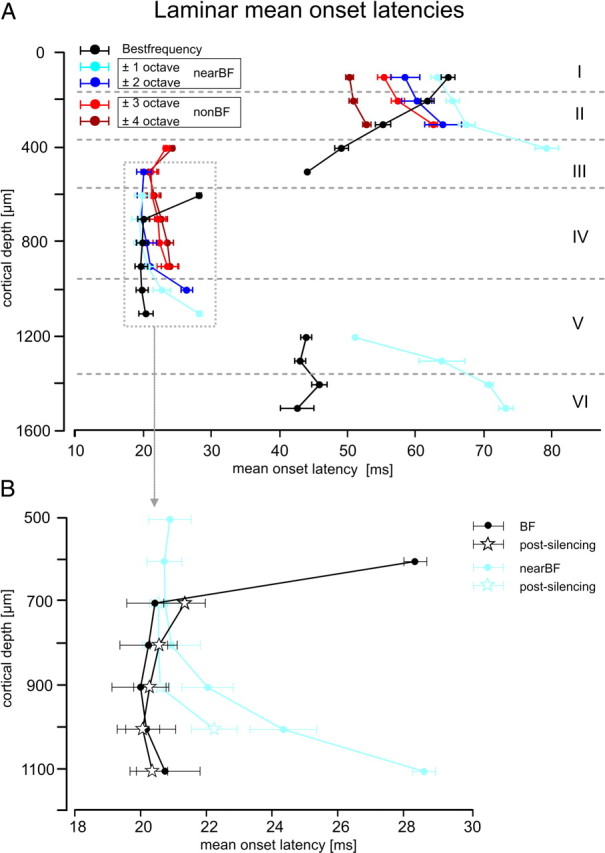
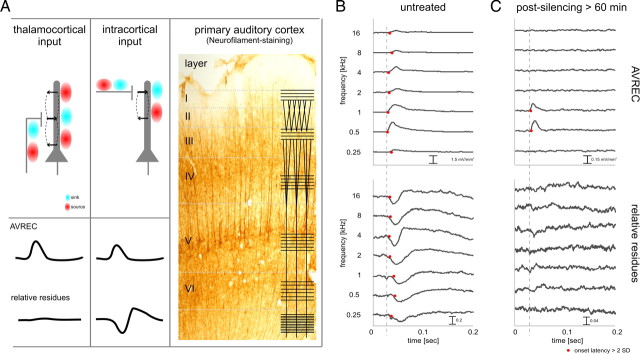
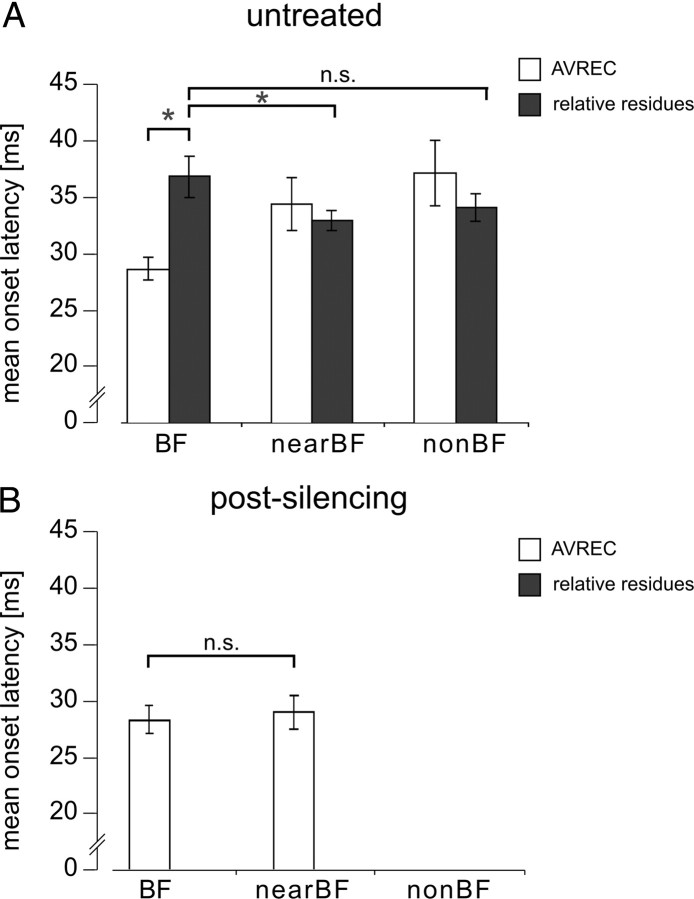
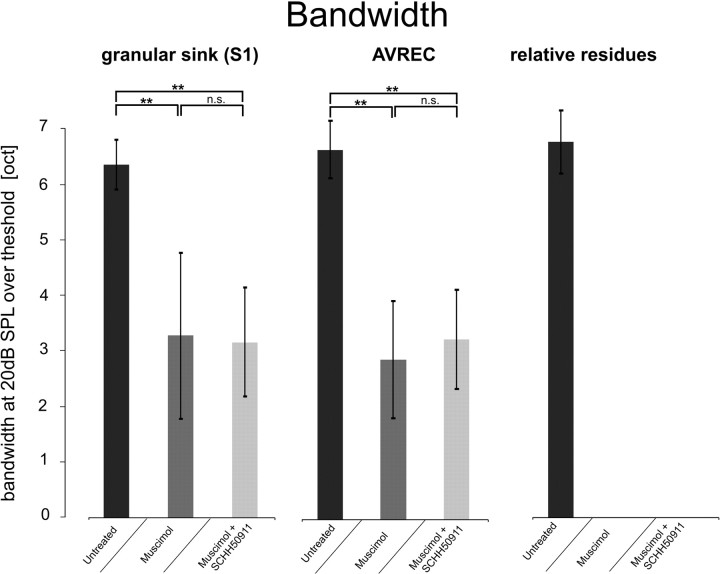
Primary sensory cortex integrates sensory information from afferent feedforward thalamocortical projection systems and convergent intracortical microcircuits. Both input systems have been demonstrated to provide different aspects of sensory information. Here we have used high-density recordings of laminar current source density (CSD) distributions in primary auditory cortex of Mongolian gerbils in combination with pharmacological silencing of cortical activity and analysis of the residual CSD, to dissociate the feedforward thalamocortical contribution and the intracortical contribution to spectral integration. We found a temporally highly precise integration of both types of inputs when the stimulation frequency was in close spectral neighborhood of the best frequency of the measurement site, in which the overlap between both inputs is maximal. Local intracortical connections provide both directly feedforward excitatory and modulatory input from adjacent cortical sites, which determine how concurrent afferent inputs are integrated. Through separate excitatory horizontal projections, terminating in cortical layers II/III, information about stimulus energy in greater spectral distance is provided even over long cortical distances. These projections effectively broaden spectral tuning width. Based on these data, we suggest a mechanism of spectral integration in primary auditory cortex that is based on temporally precise interactions of afferent thalamocortical inputs and different short- and long-range intracortical networks. The proposed conceptual framework allows integration of different and partly controversial anatomical and physiological models of spectral integration in the literature.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources