

Intrinsic circuit organization and theta-gamma oscillation dynamics in the entorhinal cortex of the rat
- PMID: 20720120
- PMCID: PMC2937273
- DOI: 10.1523/JNEUROSCI.1327-10.2010
Intrinsic circuit organization and theta-gamma oscillation dynamics in the entorhinal cortex of the rat
Abstract
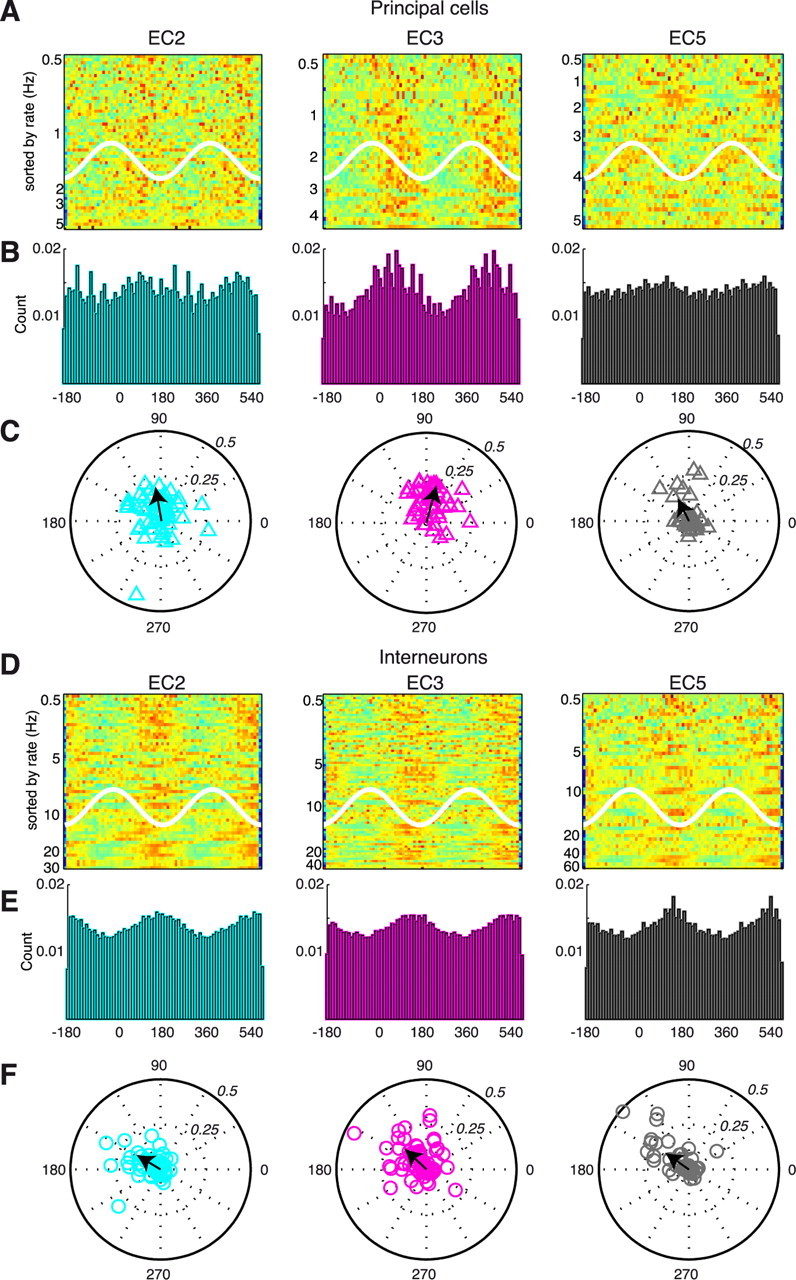
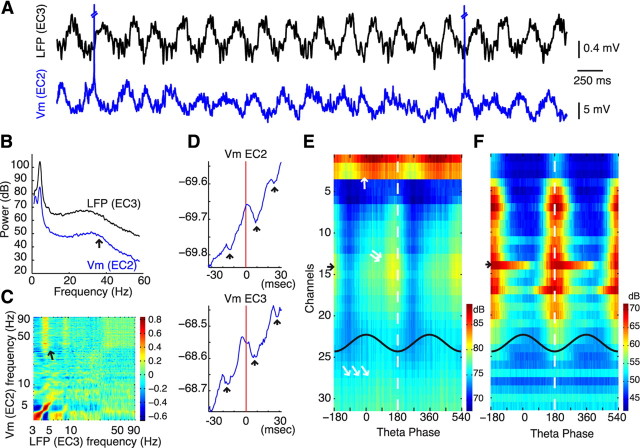
A thorough knowledge of the intrinsic circuit properties of the entorhinal cortex (EC) and the temporal dynamics these circuits support is essential for understanding how information is exchanged between the hippocampus and neocortex. Using intracellular and extracellular recordings in the anesthetized rat and anatomical reconstruction of single cells, we found that EC5 and EC2 principal neurons form large axonal networks mainly within their layers, interconnected by the more vertically organized axon trees of EC3 pyramidal cells. Principal cells showed layer-specific unique membrane properties and contributed differentially to theta and gamma oscillations. EC2 principal cells were most strongly phase modulated by EC theta. The multiple gamma oscillators, present in the various EC layers, were temporally coordinated by the phase of theta waves. Putative interneurons in all EC layers fired relatively synchronously within the theta cycle, coinciding with the maximum power of gamma oscillation. The special wiring architecture and unique membrane properties of EC neurons may underlie their behaviorally distinct firing patterns in the waking animal.
Figures









Similar articles
-
Convergence of entorhinal and CA3 inputs onto pyramidal neurons and interneurons in hippocampal area CA1--an anatomical study in the rat.Hippocampus. 2008;18(3):266-80. doi: 10.1002/hipo.20385. Hippocampus. 2008. PMID: 18000818
-
Layer-specific GABAergic control of distinct gamma oscillations in the CA1 hippocampus.Neuron. 2014 Mar 5;81(5):1126-1139. doi: 10.1016/j.neuron.2014.01.021. Neuron. 2014. PMID: 24607232
-
Gamma oscillations in the entorhinal cortex of the freely behaving rat.J Neurosci. 1998 Jan 1;18(1):388-98. doi: 10.1523/JNEUROSCI.18-01-00388.1998. J Neurosci. 1998. PMID: 9412515 Free PMC article.
-
Oscillatory activity in entorhinal neurons and circuits. Mechanisms and function.Ann N Y Acad Sci. 2000 Jun;911:127-50. doi: 10.1111/j.1749-6632.2000.tb06723.x. Ann N Y Acad Sci. 2000. PMID: 10911871 Review.
-
Consequences of parameter differences in a model of short-term persistent spiking buffers provided by pyramidal cells in entorhinal cortex.Brain Res. 2008 Apr 2;1202:54-67. doi: 10.1016/j.brainres.2007.06.067. Epub 2007 Jul 17. Brain Res. 2008. PMID: 17698043 Free PMC article. Review.
Cited by
-
Metabolic and functional connectivity changes in mal de debarquement syndrome.PLoS One. 2012;7(11):e49560. doi: 10.1371/journal.pone.0049560. Epub 2012 Nov 29. PLoS One. 2012. PMID: 23209584 Free PMC article.
-
High-frequency oscillations in human and monkey neocortex during the wake-sleep cycle.Proc Natl Acad Sci U S A. 2016 Aug 16;113(33):9363-8. doi: 10.1073/pnas.1523583113. Epub 2016 Aug 1. Proc Natl Acad Sci U S A. 2016. PMID: 27482084 Free PMC article.
-
Synaptic Mechanisms of Tight Spike Synchrony at Gamma Frequency in Cerebral Cortex.J Neurosci. 2015 Jul 15;35(28):10236-51. doi: 10.1523/JNEUROSCI.0828-15.2015. J Neurosci. 2015. PMID: 26180200 Free PMC article.
-
Architecture of spatial circuits in the hippocampal region.Philos Trans R Soc Lond B Biol Sci. 2013 Dec 23;369(1635):20120515. doi: 10.1098/rstb.2012.0515. Print 2014 Feb 5. Philos Trans R Soc Lond B Biol Sci. 2013. PMID: 24366129 Free PMC article. Review.
-
Preconfigured, skewed distribution of firing rates in the hippocampus and entorhinal cortex.Cell Rep. 2013 Sep 12;4(5):1010-21. doi: 10.1016/j.celrep.2013.07.039. Epub 2013 Aug 29. Cell Rep. 2013. PMID: 23994479 Free PMC article.
References
-
- Alonso A, García-Austt E. Neuronal sources of theta rhythm in the entorhinal cortex of the rat. I. Laminar distribution of theta field potentials. Exp Brain Res. 1987a;67:493–501. - PubMed
-
- Alonso A, García-Austt E. Neuronal sources of theta rhythm in the entorhinal cortex of the rat. II. Phase relations between unit discharges and theta field potentials. Exp Brain Res. 1987b;67:502–509. - PubMed
-
- Alonso A, Klink R. Differential electroresponsiveness of stellate and pyramidal-like cells of medial entorhinal cortex layer II. J Neurophysiol. 1993;70:128–143. - PubMed
-
- Alonso A, Llinás RR. Subthreshold Na+-dependent theta-like rhythmicity in stellate cells of entorhinal cortex layer II. Nature. 1989;342:175–177. - PubMed
-
- Amaral DG, Witter MP. The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience. 1989;31:571–591. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous