

Proximal events in 7,12-dimethylbenz[a]anthracene-induced, stromal cell-dependent bone marrow B cell apoptosis: stromal cell-B cell communication and apoptosis signaling
- PMID: 20720205
- PMCID: PMC2933279
- DOI: 10.4049/jimmunol.0902541
Proximal events in 7,12-dimethylbenz[a]anthracene-induced, stromal cell-dependent bone marrow B cell apoptosis: stromal cell-B cell communication and apoptosis signaling
Abstract
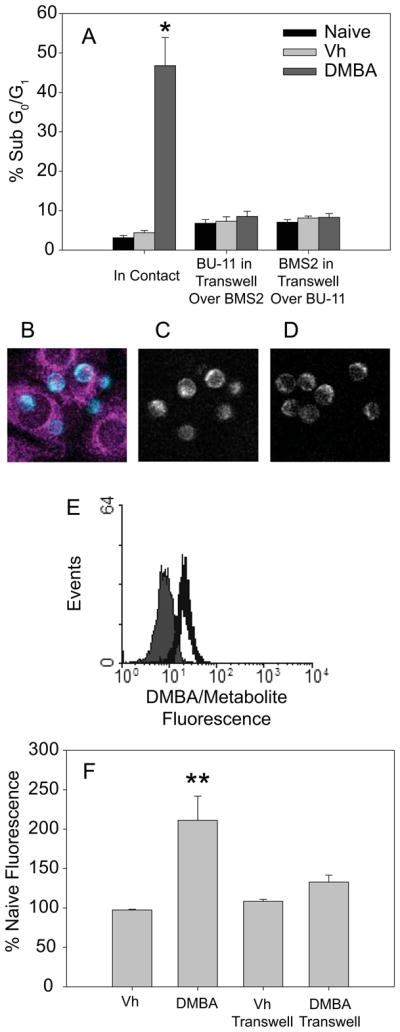
Intercellular communication is an essential process in stimulating lymphocyte development and in activating and shaping an immune response. B cell development requires cell-to-cell contact with and cytokine production by bone marrow stromal cells. However, this intimate relationship also may be responsible for the transfer of death-inducing molecules to the B cells. 7,12-Dimethylbenz[a]anthracene (DMBA), a prototypical polycyclic aromatic hydrocarbon, activates caspase-3 in pro/pre-B cells in a bone marrow stromal cell-dependent manner, resulting in apoptosis. These studies were designed to examine the hypothesis that an intrinsic apoptotic pathway is activated by DMBA and that the ultimate death signal is a DMBA metabolite generated by the stromal cells and transferred to the B cells. Although a loss of mitochondrial membrane potential did not occur in the DMBA/stromal cell-induced pathway, cytochrome c release was stimulated in B cells. Caspase-9 was activated, and formation of the apoptosome was required to support apoptosis, as demonstrated by the suppression of death in Apaf-1(fog) mutant pro-B cells. Investigation of signaling upstream of the mitochondria demonstrated an essential role for p53. Furthermore, DMBA-3,4-dihydrodiol-1,2-epoxide, a DNA-reactive metabolite of DMBA, was sufficient to upregulate p53, induce caspase-9 cleavage, and initiate B cell apoptosis in the absence of stromal cells, suggesting that production of this metabolite by the stromal cells and transfer to the B cells are proximal events in triggering apoptosis. Indeed, we provide evidence that metabolite transfer from bone marrow stromal cells occurs through membrane exchange, which may represent a novel communication mechanism between developing B cells and stromal cells.
Figures





References
-
- White KL, Jr, Lysy HH, Holsapple MP. Immunosuppression by polycyclic aromatic hydrocarbons: A structure-activity relationship in B6C3F1 and DBA/2 mice. Immunopharmacology. 1985;9:155–164. - PubMed
-
- Dean J, Ward E, Murray M, Lauer L, House R, Stillman W, Hamilton T, Adams D. Immunosuppression following 7,12-deimthylbenz[a]anthracene exposure in B6C3F1--II. Altered cell-mediated immunity and tumore resistance. Int J Immunopharm. 1986;8:189–198. - PubMed
-
- Thurmond L, Lauer L, House R, Cook J, Dean J. Immunosuppression following exposure to 7,12-dimethylbenz[a]anthracene (DMBA) in Ah-responsive and Ah-nonresponsive mice. Toxicology and Applied Pharmacology. 1987;91:450–460. - PubMed
-
- Ward EC, Murray MJ, Lauer LD, House RV, Irons R, Dean JH. Immunosuppression following 7,12-dimethylbenz[a]anthracene exposure in B6C3F1 mice. I. Effect on humoral immunity and host resistance. Toxicol Appl Pharmacol. 1984;75:299–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
