

The monopolin complex crosslinks kinetochore components to regulate chromosome-microtubule attachments
- PMID: 20723757
- PMCID: PMC2955198
- DOI: 10.1016/j.cell.2010.07.017
The monopolin complex crosslinks kinetochore components to regulate chromosome-microtubule attachments
Abstract
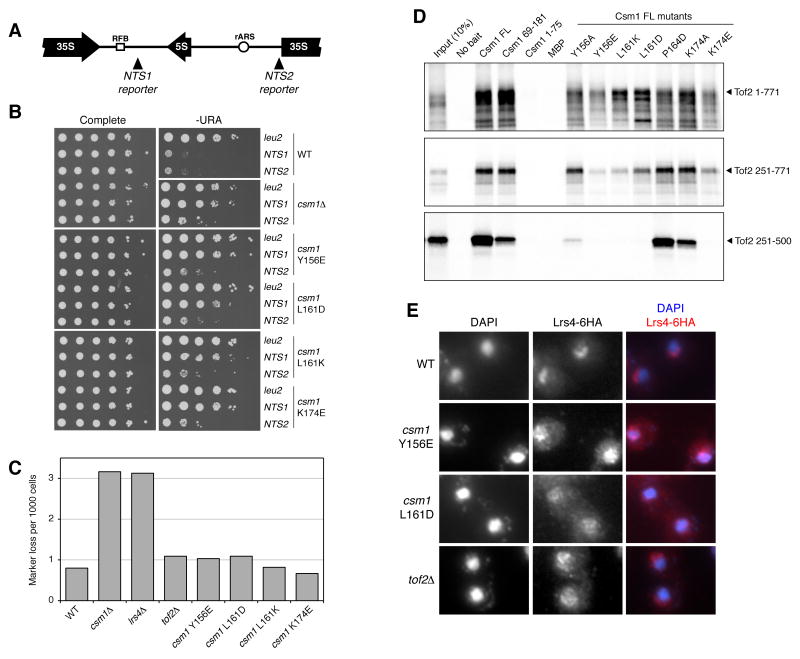
The monopolin complex regulates different types of kinetochore-microtubule attachments in fungi, ensuring sister chromatid co-orientation in Saccharomyces cerevisiae meiosis I and inhibiting merotelic attachment in Schizosaccharomyces pombe mitosis. In addition, the monopolin complex maintains the integrity and silencing of ribosomal DNA (rDNA) repeats in the nucleolus. We show here that the S. cerevisiae Csm1/Lrs4 monopolin subcomplex has a distinctive V-shaped structure, with two pairs of protein-protein interaction domains positioned approximately 10 nm apart. Csm1 presents a conserved hydrophobic surface patch that binds two kinetochore proteins: Dsn1, a subunit of the outer-kinetochore MIND/Mis12 complex, and Mif2/CENP-C. Csm1 point-mutations that disrupt kinetochore-subunit binding also disrupt sister chromatid co-orientation in S. cerevisiae meiosis I. We further show that the same Csm1 point-mutations affect rDNA silencing, probably by disrupting binding to the rDNA-associated protein Tof2. We propose that Csm1/Lrs4 functions as a molecular clamp, crosslinking kinetochore components to enforce sister chromatid co-orientation in S. cerevisiae meiosis I and to suppress merotelic attachment in S. pombe mitosis, and crosslinking rDNA repeats to aid rDNA silencing.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures





Similar articles
-
The molecular basis of monopolin recruitment to the kinetochore.Chromosoma. 2019 Sep;128(3):331-354. doi: 10.1007/s00412-019-00700-0. Epub 2019 Apr 30. Chromosoma. 2019. PMID: 31037469 Free PMC article.
-
Monopolin subunit Csm1 associates with MIND complex to establish monopolar attachment of sister kinetochores at meiosis I.PLoS Genet. 2013;9(7):e1003610. doi: 10.1371/journal.pgen.1003610. Epub 2013 Jul 4. PLoS Genet. 2013. PMID: 23861669 Free PMC article.
-
Kinetochore recruitment of two nucleolar proteins is required for homolog segregation in meiosis I.Dev Cell. 2003 Apr;4(4):535-48. doi: 10.1016/s1534-5807(03)00086-8. Dev Cell. 2003. PMID: 12689592
-
Studies of meiosis disclose distinct roles of cohesion in the core centromere and pericentromeric regions.Chromosome Res. 2009;17(2):239-49. doi: 10.1007/s10577-008-9013-y. Chromosome Res. 2009. PMID: 19308704 Review.
-
Kinetochore composition and its function: lessons from yeasts.FEMS Microbiol Rev. 2014 Mar;38(2):185-200. doi: 10.1111/1574-6976.12049. FEMS Microbiol Rev. 2014. PMID: 24666101 Review.
Cited by
-
Meiosis in male Drosophila.Spermatogenesis. 2012 Jul 1;2(3):167-184. doi: 10.4161/spmg.21800. Spermatogenesis. 2012. PMID: 23087836 Free PMC article.
-
Regional centromere configuration in the fungal pathogens of the Pneumocystis genus.mBio. 2024 Mar 13;15(3):e0318523. doi: 10.1128/mbio.03185-23. Epub 2024 Feb 21. mBio. 2024. PMID: 38380929 Free PMC article.
-
Acetylation of the SUN protein Mps3 by Eco1 regulates its function in nuclear organization.Mol Biol Cell. 2012 Jul;23(13):2546-59. doi: 10.1091/mbc.E11-07-0600. Epub 2012 May 16. Mol Biol Cell. 2012. PMID: 22593213 Free PMC article.
-
Meiosis: an overview of key differences from mitosis.Cold Spring Harb Perspect Biol. 2015 Jan 20;7(5):a015859. doi: 10.1101/cshperspect.a015859. Cold Spring Harb Perspect Biol. 2015. PMID: 25605710 Free PMC article. Review.
-
Establishing correct kinetochore-microtubule attachments in mitosis and meiosis.Essays Biochem. 2020 Sep 4;64(2):277-287. doi: 10.1042/EBC20190072. Essays Biochem. 2020. PMID: 32406497 Free PMC article. Review.
References
-
- Cheeseman IM, Chappie JS, Wilson-Kubalek EM, Desai A. The conserved KMN network constitutes the core microtubule-binding site of the kinetochore. Cell. 2006;127:983–997. - PubMed
-
- Cheeseman IM, Desai A. Molecular architecture of the kinetochore-microtubule interface. Nat Rev Mol Cell Biol. 2008;9:33–46. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
