

Structural basis for negative cooperativity in growth factor binding to an EGF receptor
- PMID: 20723758
- PMCID: PMC2925043
- DOI: 10.1016/j.cell.2010.07.015
Structural basis for negative cooperativity in growth factor binding to an EGF receptor
Abstract
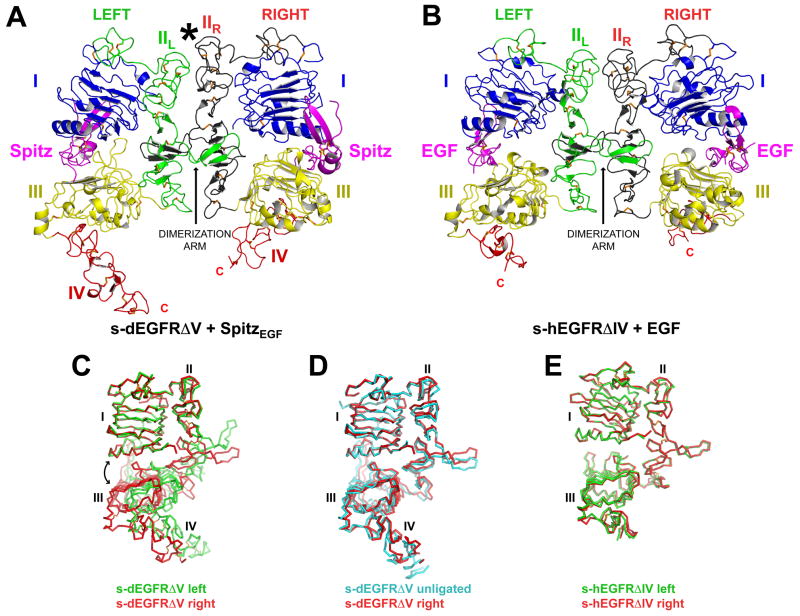
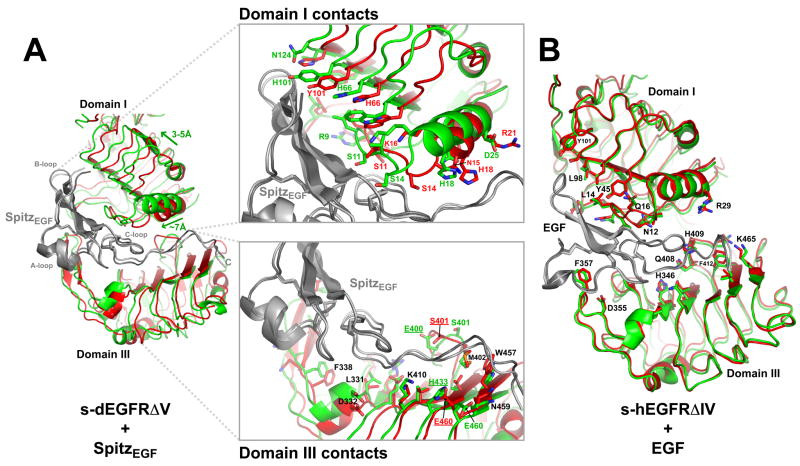
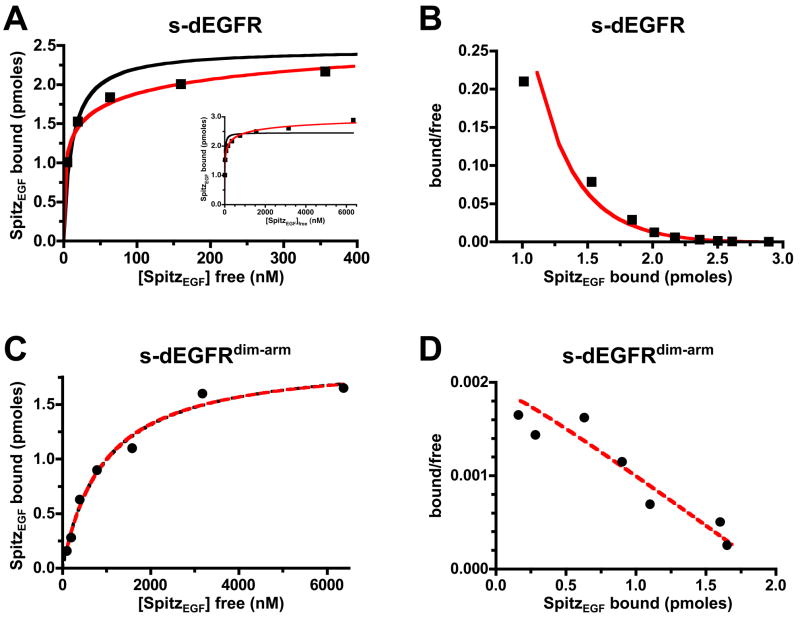
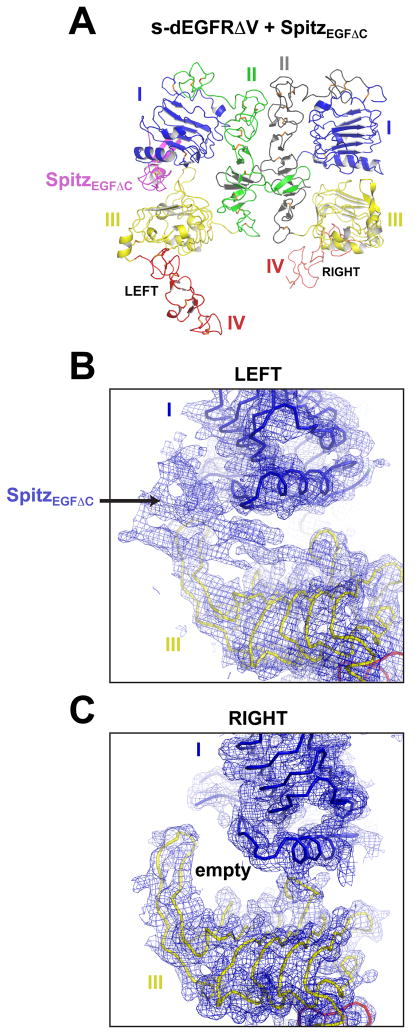
Transmembrane signaling by the epidermal growth factor receptor (EGFR) involves ligand-induced dimerization and allosteric regulation of the intracellular tyrosine kinase domain. Crystallographic studies have shown how ligand binding induces dimerization of the EGFR extracellular region but cannot explain the "high-affinity" and "low-affinity" classes of cell-surface EGF-binding sites inferred from curved Scatchard plots. From a series of crystal structures of the Drosophila EGFR extracellular region, we show here how Scatchard plot curvature arises from negatively cooperative ligand binding. The first ligand-binding event induces formation of an asymmetric dimer with only one bound ligand. The unoccupied site in this dimer is structurally restrained, leading to reduced affinity for binding of the second ligand, and thus negative cooperativity. Our results explain the cell-surface binding characteristics of EGF receptors and suggest how individual EGFR ligands might stabilize distinct dimeric species with different signaling properties.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
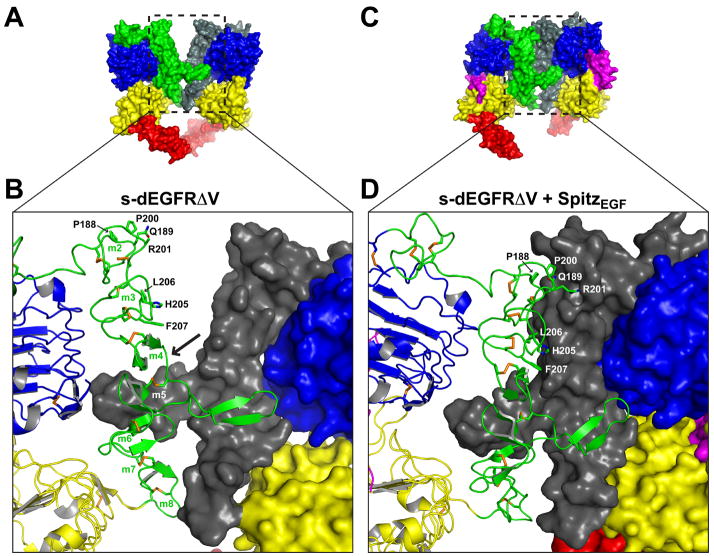
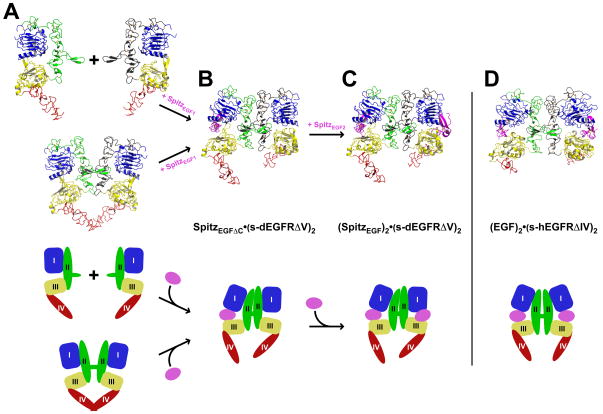
References
-
- Adams TE, Koziolek EJ, Hoyne PH, Bentley JD, Lu L, Lovrecz G, Ward CW, Lee FT, Scott AM, Nash AD, et al. A truncated soluble epidermal growth factor receptor-Fc fusion ligand trap displays anti-tumour activity in vivo. Growth Factors. 2009;27:141–154. - PubMed
-
- Bass J, Kurose T, Pashmforoush M, Steiner DF. Fusion of insulin receptor ectodomains to immunoglobulin constant domains reproduces high-affinity insulin binding in vitro. J Biol Chem. 1996;271:19367–19375. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
