

Activation of specific apoptotic caspases with an engineered small-molecule-activated protease
- PMID: 20723762
- PMCID: PMC3689538
- DOI: 10.1016/j.cell.2010.07.014
Activation of specific apoptotic caspases with an engineered small-molecule-activated protease
Abstract
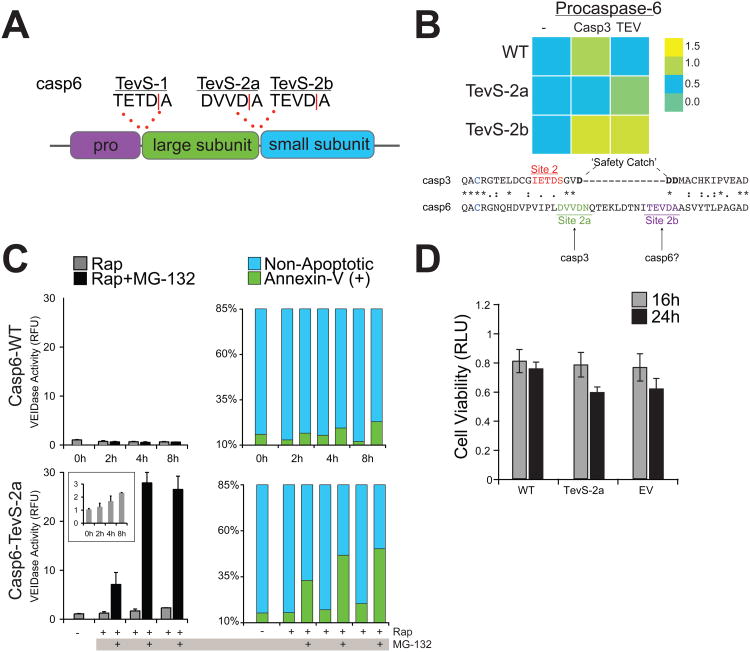
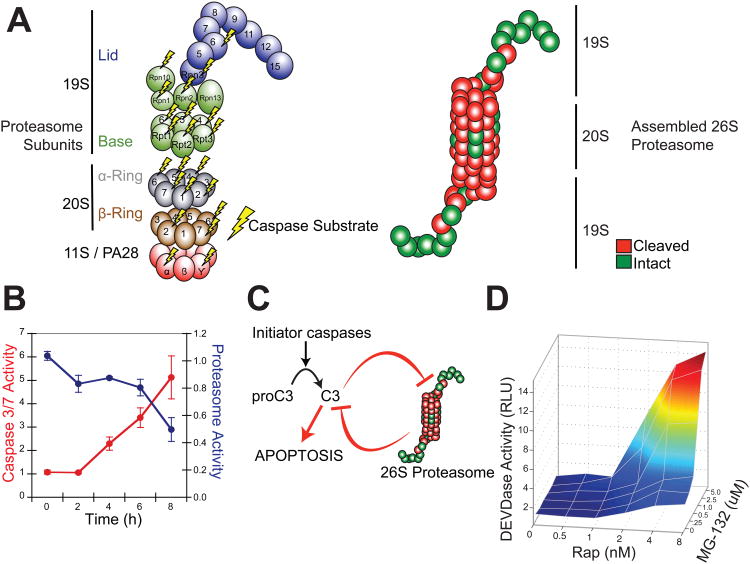
Apoptosis is a conserved cellular pathway that results in the activation of cysteine-aspartyl proteases, or caspases. To dissect the nonredundant roles of the executioner caspase-3, -6, and -7 in orchestrating apoptosis, we have developed an orthogonal protease to selectively activate each isoform in human cells. Our approach uses a split-tobacco etch virus (TEV) protease under small-molecule control, which we call the SNIPer, with caspase alleles containing genetically encoded TEV cleavage sites. These studies reveal that all three caspases are transiently activated but only activation of caspase-3 or -7 is sufficient to induce apoptosis. Proteomic analysis shown here and from others reveals that 20 of the 33 subunits of the 26S proteasome can be cut by caspases, and we demonstrate synergy between proteasome inhibition and dose-dependent caspase activation. We propose a model of proteolytic reciprocal negative regulation with mechanistic implications for the combined clinical use of proteasome inhibitors and proapoptotic drugs.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
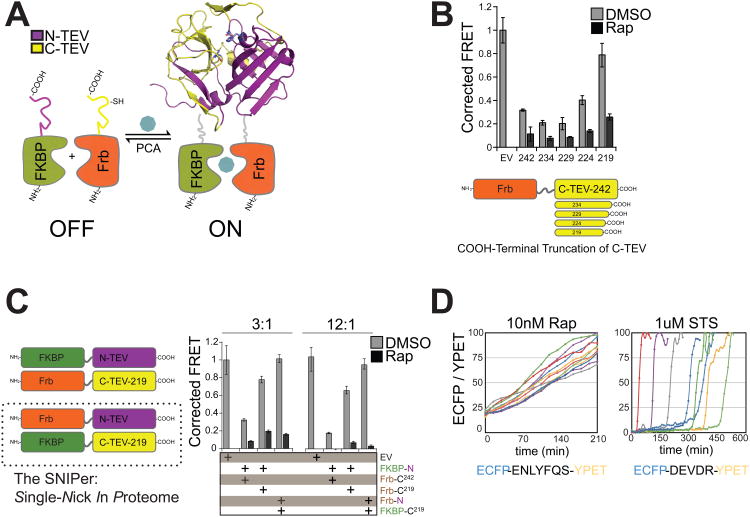
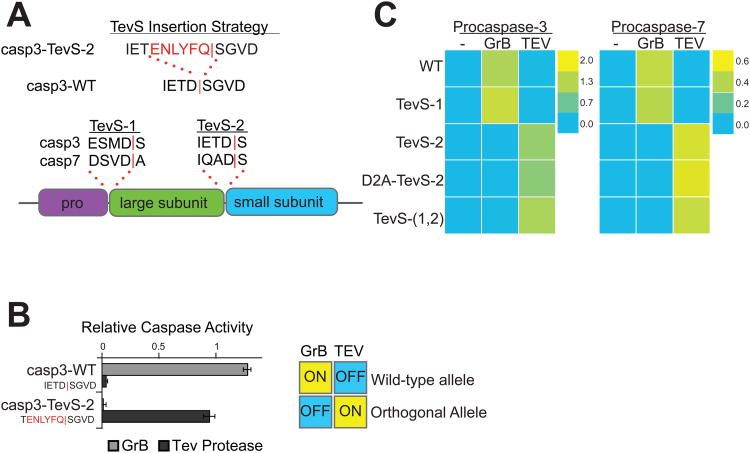
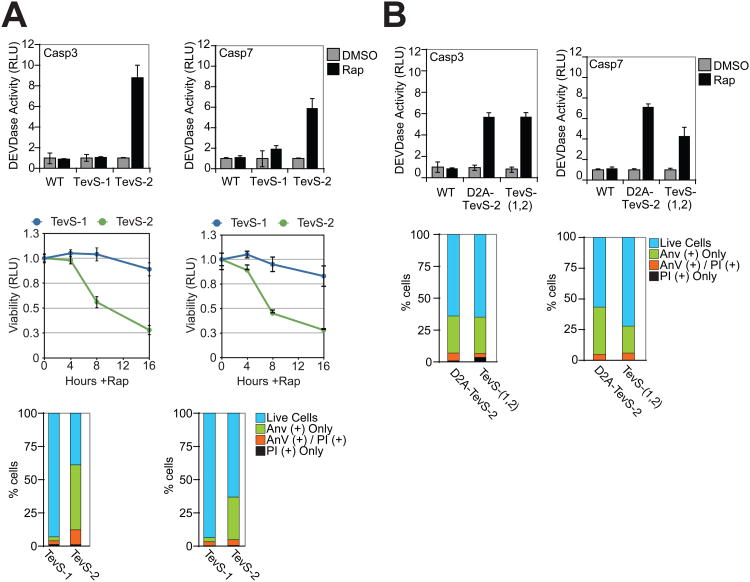
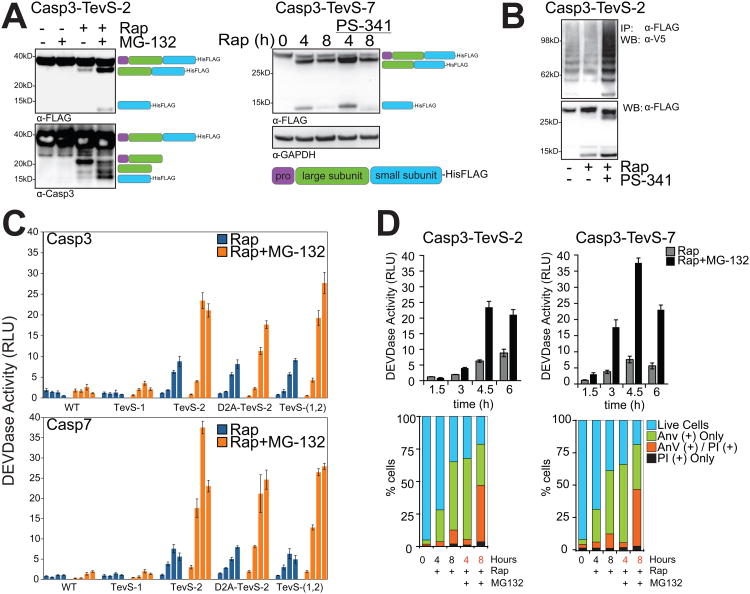
Comment in
-
SNIPer pulls the trigger.Nat Methods. 2010 Oct;7(10):786. doi: 10.1038/nmeth1010-786. Nat Methods. 2010. PMID: 20936776 No abstract available.
References
-
- Adrain C, Creagh EM, Cullen SP, Martin SJ. Caspase-dependent inactivation of proteasome function during programmed cell death in Drosophila and man. J Biol Chem. 2004;279:36923–36930. - PubMed
-
- Casciola-Rosen L, Garcia-Calvo M, Bull H, Becker J, Hines T, Thornberry N, Rosen A. Mouse and Human Granzyme B Have Distinct Tetrapeptide Specificities and Abilities to Recruit the Bid Pathway. Journal of Biological Chemistry. 2007;282:4545. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
