

The malondialdehyde-derived fluorophore DHP-lysine is a potent sensitizer of UVA-induced photooxidative stress in human skin cells
- PMID: 20724175
- PMCID: PMC2963670
- DOI: 10.1016/j.jphotobiol.2010.07.010
The malondialdehyde-derived fluorophore DHP-lysine is a potent sensitizer of UVA-induced photooxidative stress in human skin cells
Abstract
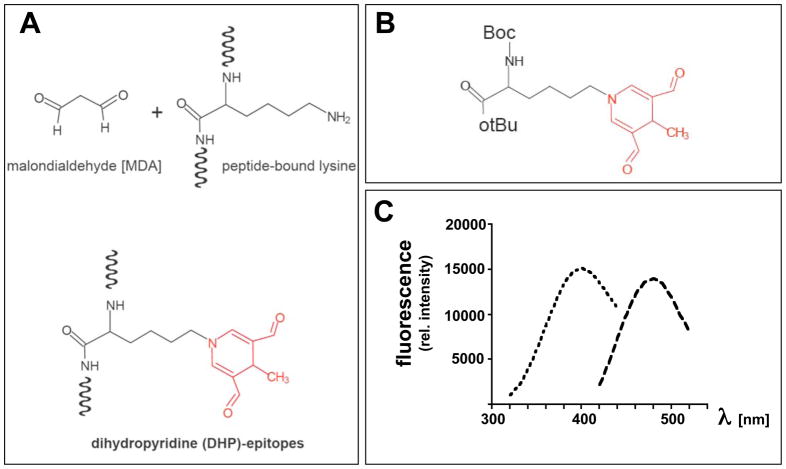
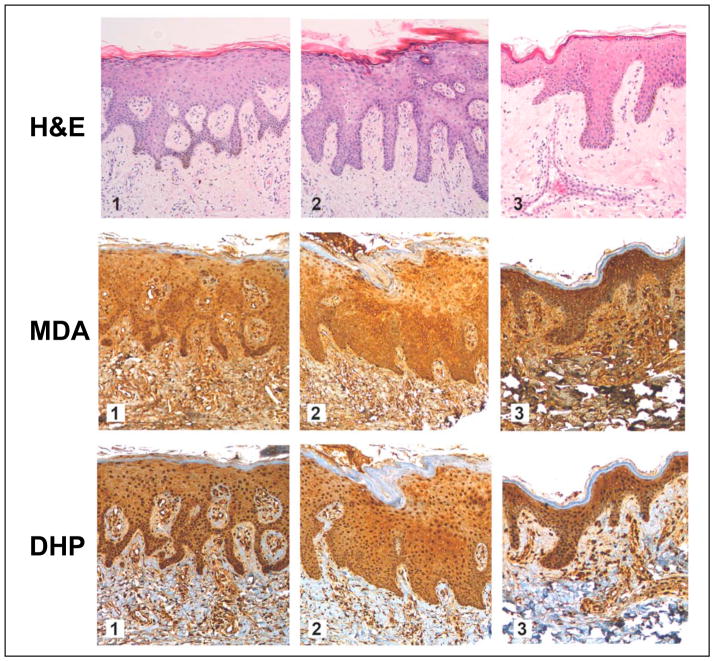
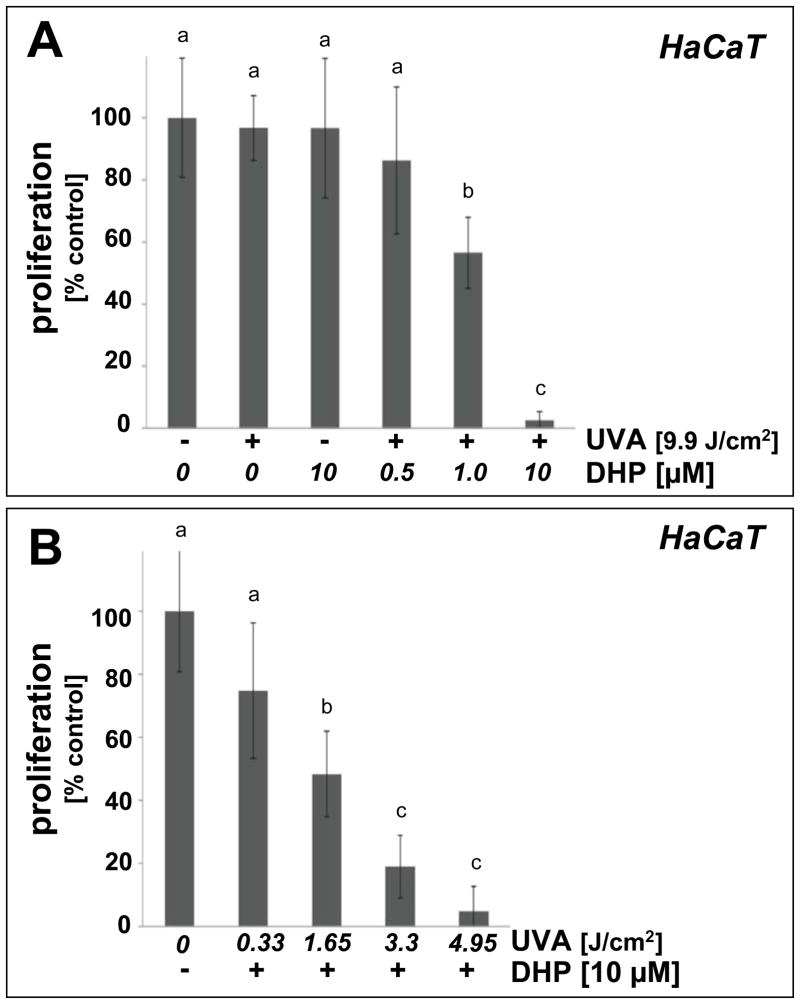
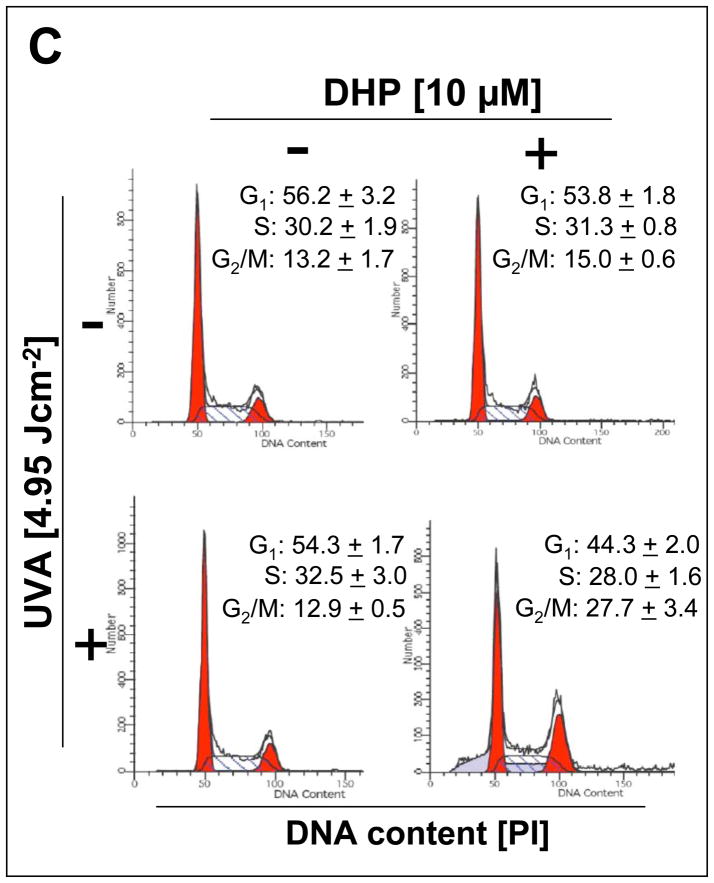
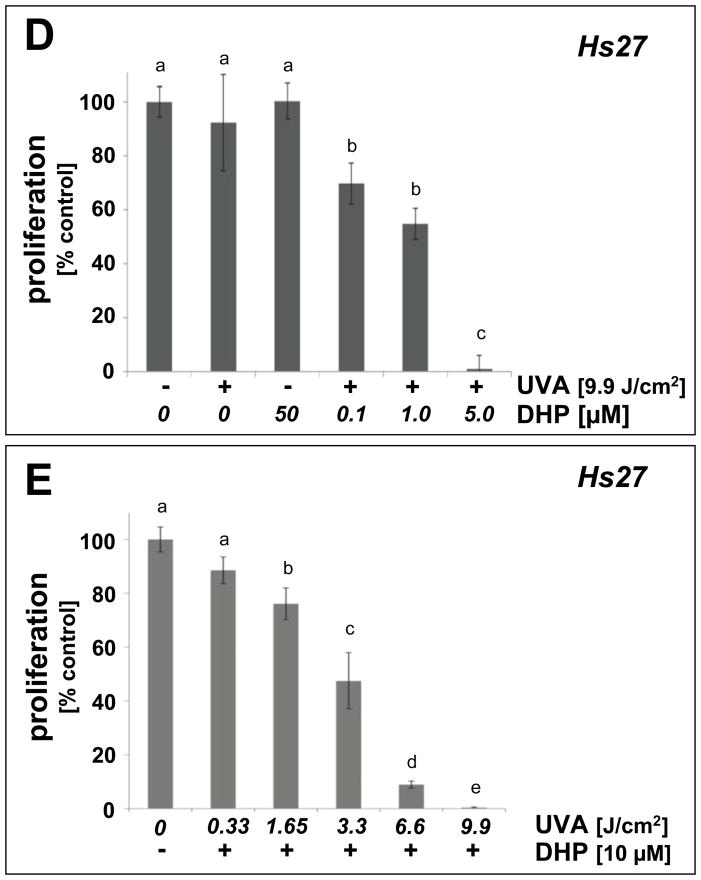
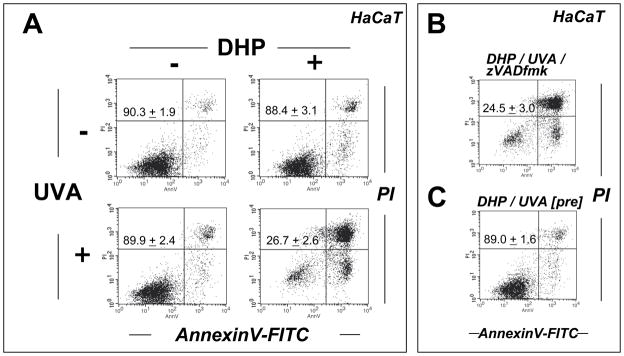
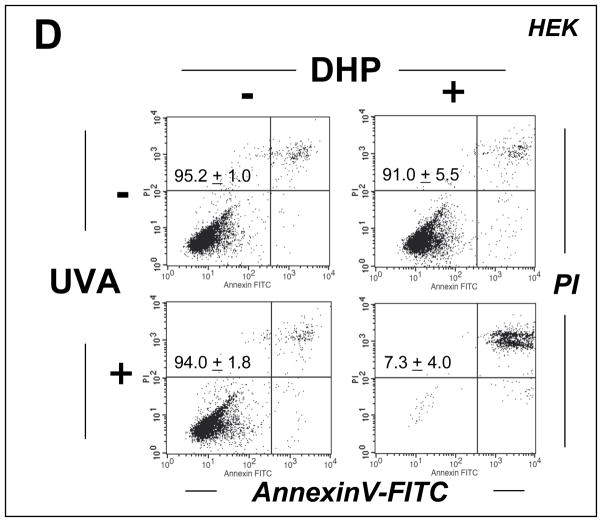
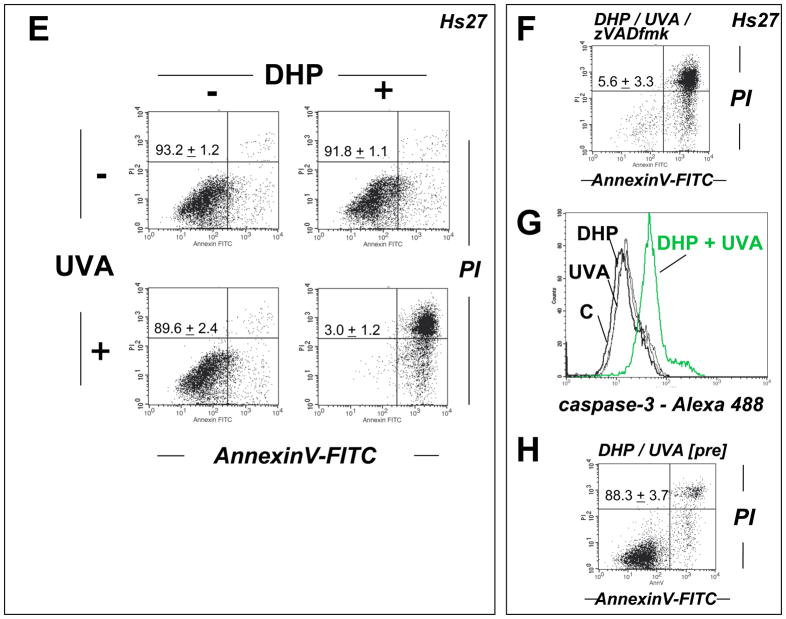
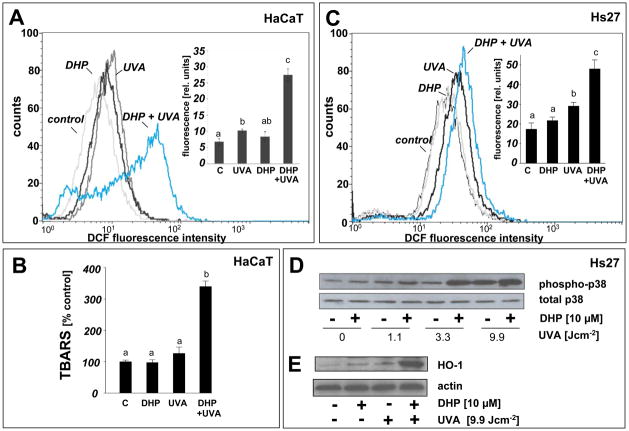
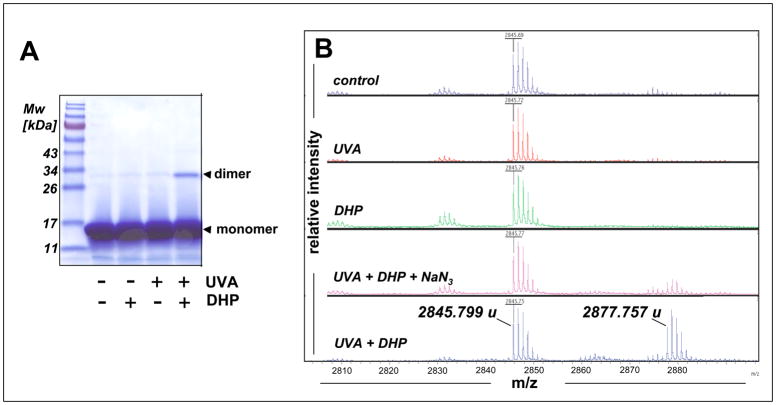
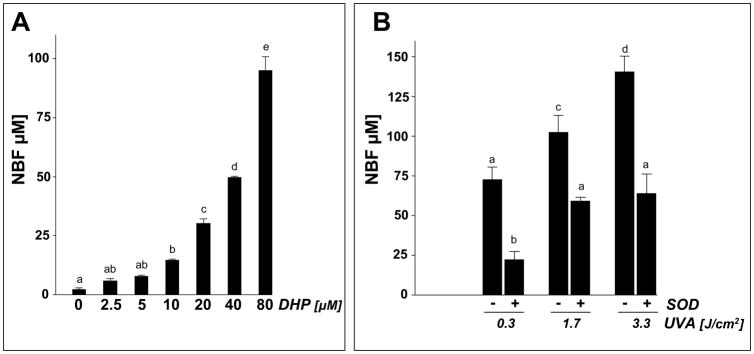
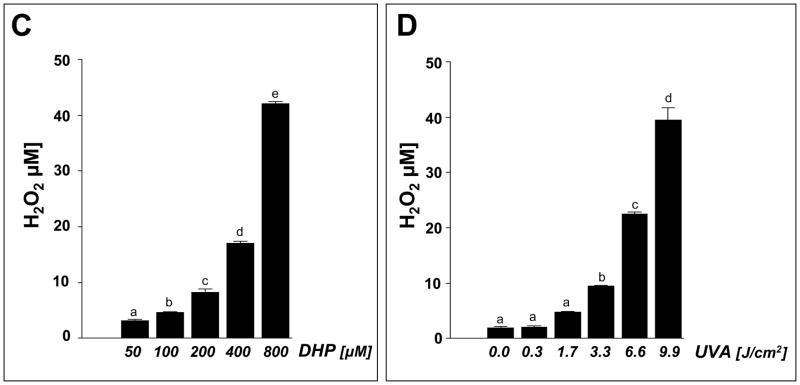
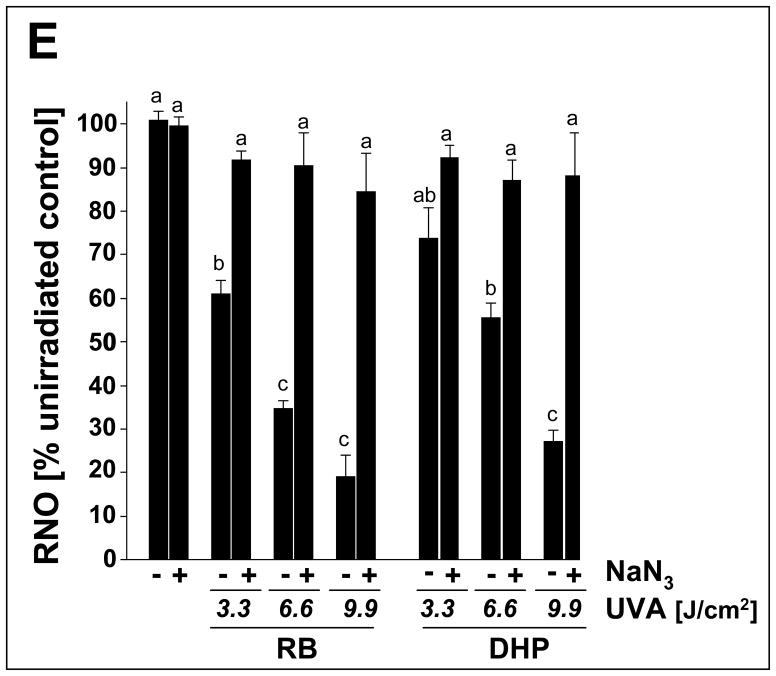
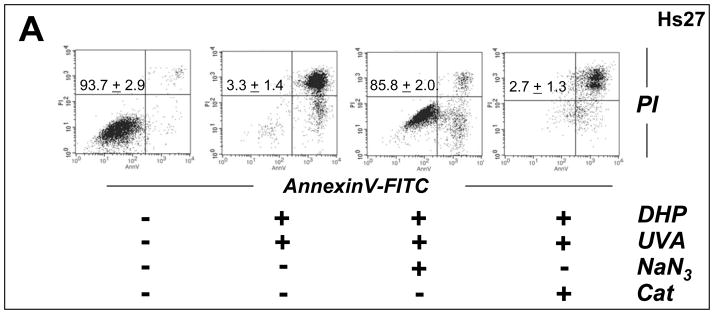
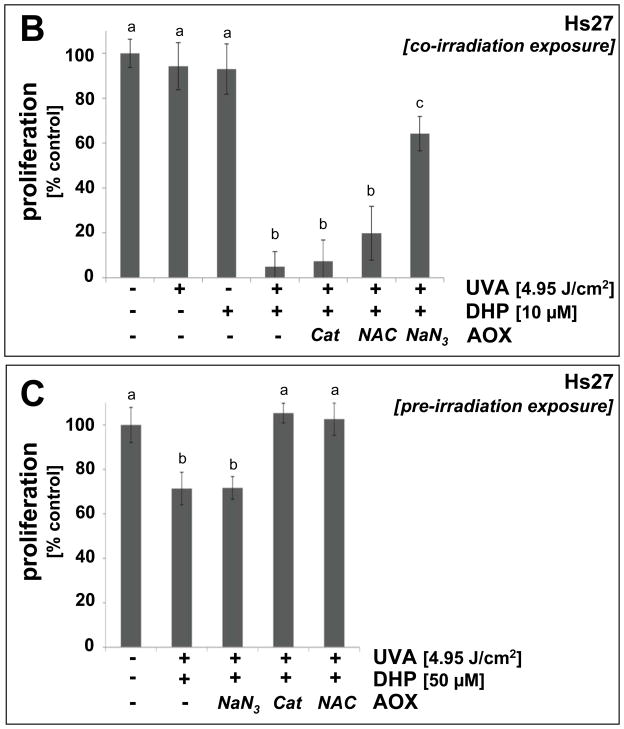
Light-driven electron and energy transfer involving non-DNA skin chromophores as endogenous photosensitizers induces oxidative stress in UVA-exposed human skin, a process relevant to photoaging and photocarcinogenesis. Malondialdehyde is an electrophilic dicarbonyl-species derived from membrane lipid peroxidation. Here, we present experimental evidence suggesting that the malondialdehyde-derived protein epitope dihydropyridine (DHP)-lysine is a potent endogenous UVA-photosensitizer of human skin cells. Immunohistochemical analysis revealed the abundant occurrence of malondialdehyde-derived and DHP-lysine epitopes in human skin. Using the chemically protected dihydropyridine-derivative (2S)-Boc-2-amino-6-(3,5-diformyl-4-methyl-4H-pyridin-1-yl)-hexanoic acid-t-butylester as a model of peptide-bound DHP-lysine, photodynamic inhibition of proliferation and induction of cell death were observed in human skin Hs27 fibroblasts as well as primary and HaCaT keratinocytes exposed to the combined action of UVA and DHP-lysine. DHP-lysine photosensitization induced intracellular oxidative stress, p38 MAPkinase activation, and upregulation of heme oxygenase-1 expression. Consistent with UVA-driven ROS formation from DHP-lysine, formation of superoxide, hydrogen peroxide, and singlet oxygen was detected in chemical assays, but little protection was achieved using SOD or catalase during cellular photosensitization. In contrast, inclusion of NaN(3) completely abolished DHP-photosensitization. Taken together, these data demonstrate photodynamic activity of DHP-lysine and support the hypothesis that malondialdehyde-derived protein-epitopes may function as endogenous sensitizers of UVA-induced oxidative stress in human skin.
Copyright © 2010 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Malondialdehyde-derived epitopes in human skin result from acute exposure to solar UV and occur in nonmelanoma skin cancer tissue.J Photochem Photobiol B. 2014 Mar 5;132:56-65. doi: 10.1016/j.jphotobiol.2014.01.019. Epub 2014 Feb 12. J Photochem Photobiol B. 2014. PMID: 24584085 Free PMC article.
-
3-hydroxypyridine chromophores are endogenous sensitizers of photooxidative stress in human skin cells.J Biol Chem. 2004 Jul 16;279(29):30009-20. doi: 10.1074/jbc.M404379200. Epub 2004 May 7. J Biol Chem. 2004. PMID: 15133022
-
The Tryptophan-Derived Endogenous Aryl Hydrocarbon Receptor Ligand 6-Formylindolo[3,2-b]Carbazole Is a Nanomolar UVA Photosensitizer in Epidermal Keratinocytes.J Invest Dermatol. 2015 Jun;135(6):1649-1658. doi: 10.1038/jid.2014.503. Epub 2014 Nov 28. J Invest Dermatol. 2015. PMID: 25431849 Free PMC article.
-
High efficiency of 5-aminolevulinate-photodynamic treatment using UVA irradiation.Carcinogenesis. 2001 Jun;22(6):879-83. doi: 10.1093/carcin/22.6.879. Carcinogenesis. 2001. PMID: 11375893
-
Endogenous UVA-photosensitizers: mediators of skin photodamage and novel targets for skin photoprotection.Photochem Photobiol Sci. 2006 Feb;5(2):215-37. doi: 10.1039/b504573h. Epub 2005 Aug 19. Photochem Photobiol Sci. 2006. PMID: 16465308 Review.
Cited by
-
The Nrf2-inducers tanshinone I and dihydrotanshinone protect human skin cells and reconstructed human skin against solar simulated UV.Redox Biol. 2013 Oct 29;1(1):532-41. doi: 10.1016/j.redox.2013.10.004. eCollection 2013. Redox Biol. 2013. PMID: 24273736 Free PMC article.
-
High-Density Lipoprotein (HDL) Triglyceride and Oxidized HDL: New Lipid Biomarkers of Lipoprotein-Related Atherosclerotic Cardiovascular Disease.Antioxidants (Basel). 2020 Apr 26;9(5):362. doi: 10.3390/antiox9050362. Antioxidants (Basel). 2020. PMID: 32357465 Free PMC article.
-
Primary Processes of Free Radical Formation in Pharmaceutical Formulations of Therapeutic Proteins.Biomolecules. 2023 Jul 17;13(7):1142. doi: 10.3390/biom13071142. Biomolecules. 2023. PMID: 37509177 Free PMC article. Review.
-
Autophagic-lysosomal dysregulation downstream of cathepsin B inactivation in human skin fibroblasts exposed to UVA.Photochem Photobiol Sci. 2012 Jan;11(1):163-72. doi: 10.1039/c1pp05131h. Epub 2011 Jul 20. Photochem Photobiol Sci. 2012. PMID: 21773629 Free PMC article.
-
Photo-Degradation of Therapeutic Proteins: Mechanistic Aspects.Pharm Res. 2020 Feb 3;37(3):45. doi: 10.1007/s11095-020-2763-8. Pharm Res. 2020. PMID: 32016661 Review.
References
-
- Scharffetter-Kochanek K, Wlaschek M, Brenneisen P, Schauen M, Blaudschun R, Wenk J. UV-induced reactive oxygen species in photocarcinogenesis and photoaging. Biol Chem. 1997;378:1247–1257. - PubMed
-
- Wondrak GT, Jacobson MK, Jacobson EL. Endogenous UVA-photosensitizers: mediators of skin photodamage and novel targets for skin photoprotection. Photochem Photobiol Sci. 2006;5:215–237. - PubMed
-
- Halliday GM. Inflammation, gene mutation and photoimmunosuppression in response to UVR-induced oxidative damage contributes to photocarcinogenesis. Mutat Res. 2005;571:107–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
