

Identification, subcellular localization, biochemical properties, and high-resolution crystal structure of Trypanosoma brucei UDP-glucose pyrophosphorylase
- PMID: 20724435
- PMCID: PMC3270307
- DOI: 10.1093/glycob/cwq115
Identification, subcellular localization, biochemical properties, and high-resolution crystal structure of Trypanosoma brucei UDP-glucose pyrophosphorylase
Abstract
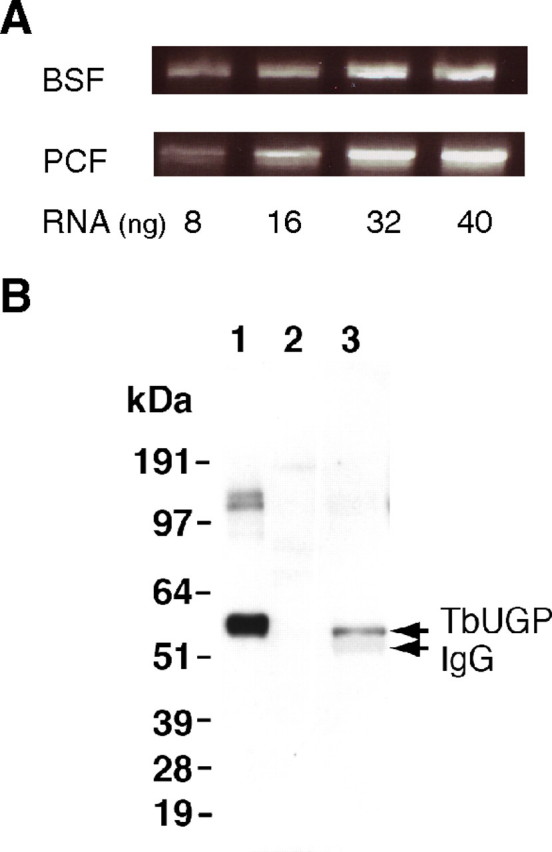
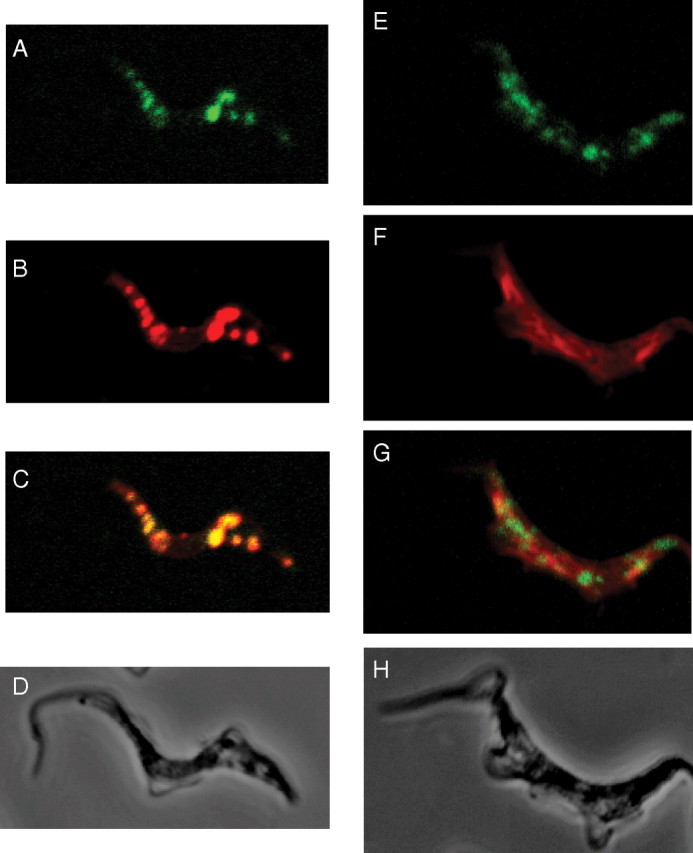
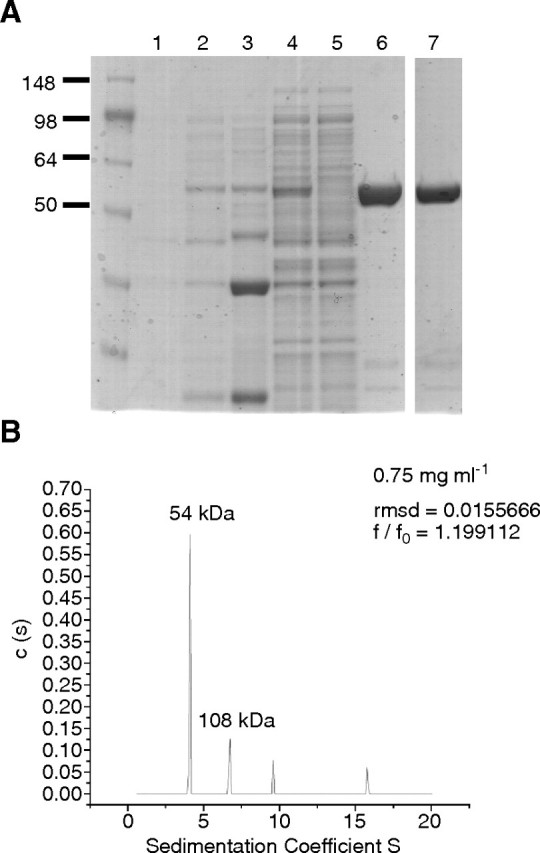
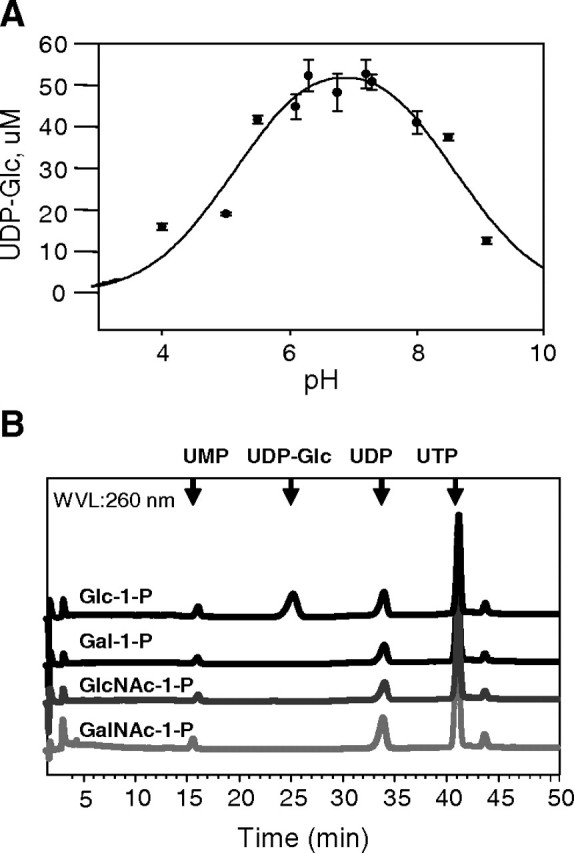
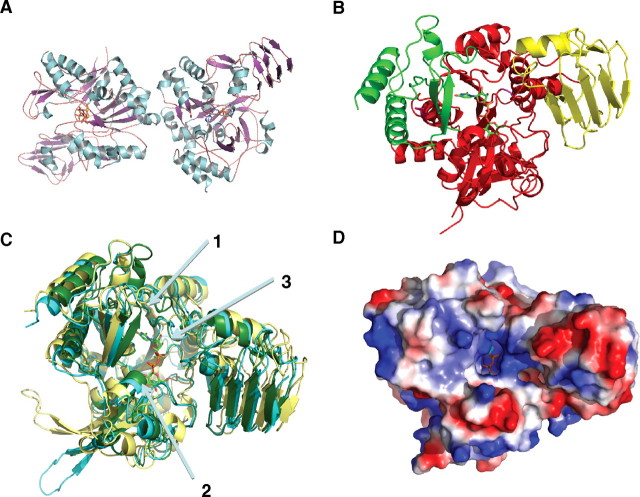
The protozoan parasite Trypanosoma brucei is the causative agent of the cattle disease Nagana and human African sleeping sickness. Glycoproteins play key roles in the parasite's survival and infectivity, and the de novo biosyntheses of the sugar nucleotides UDP-galactose (UDP-Gal), UDP-N-acetylglucosamine, and GDP-fucose have been shown to be essential for their growth. The only route to UDP-Gal in T. brucei is through the epimerization of UDP-glucose (UDP-Glc) by UDP-Glc 4'-epimerase. UDP-Glc is also the glucosyl donor for the unfolded glycoprotein glucosyltransferase (UGGT) involved in glycoprotein quality control in the endoplasmic reticulum and is the presumed donor for the synthesis of base J (β-D-glucosylhydroxymethyluracil), a rare deoxynucleotide found in telomere-proximal DNA in the bloodstream form of T. brucei. Considering that UDP-Glc plays such a central role in carbohydrate metabolism, we decided to characterize UDP-Glc biosynthesis in T. brucei. We identified and characterized the parasite UDP-glucose pyrophosphorylase (TbUGP), responsible for the formation of UDP-Glc from glucose-1-phosphate and UTP, and localized the enzyme to the peroxisome-like glycosome organelles of the parasite. Recombinant TbUGP was shown to be enzymatically active and specific for glucose-1-phosphate. The high-resolution crystal structure was also solved, providing a framework for the design of potential inhibitors against the parasite enzyme.
Figures

References
-
- Alexander DL, Schwartz KJ, Balber AE, Bangs JD. Developmentally regulated trafficking of the lysosomal membrane protein p67 in Trypanosoma brucei. J Cell Sci. 2002;115:3253–3263. - PubMed
-
- Chang H-Y, Peng H-L, Chao YC, Duggleby RG. The importance of conserved residues in human liver UDP-glucose pyrophosphorylase. Eur J Biochem. 1996;236:723–728. - PubMed
-
- Chen R, Zhao X, Shao Z, Zhu L, He G. Multiple isoforms of UDP-glucose pyrophosphorylase in rice. Physiol Plant. 2007;129:725–736.
-
- Collaborative Computational Project, Number 4. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr D Biol Crystallogr. 1994;50:760–763. - PubMed
-
- Cross GA. Antigenic variation in trypanosomes: Secrets surface slowly. Bioessays. 1996;18:283–291. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
