

Regulation of NADPH oxidase activity in phagocytes: relationship between FAD/NADPH binding and oxidase complex assembly
- PMID: 20724480
- PMCID: PMC2963400
- DOI: 10.1074/jbc.M110.151555
Regulation of NADPH oxidase activity in phagocytes: relationship between FAD/NADPH binding and oxidase complex assembly
Abstract
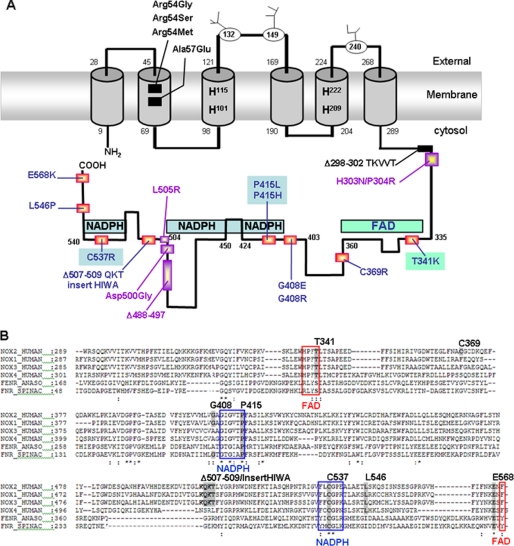
The X(+)-linked chronic granulomatous disease (X(+)-CGD) variants are natural mutants characterized by defective NADPH oxidase activity but with normal Nox2 expression. According to the three-dimensional model of the cytosolic Nox2 domain, most of the X(+)-CGD mutations are located in/or close to the FAD/NADPH binding regions. A structure/function study of this domain was conducted in X(+)-CGD PLB-985 cells exactly mimicking 10 human variants: T341K, C369R, G408E, G408R, P415H, P415L, Δ507QKT509-HIWAinsert, C537R, L546P, and E568K. Diaphorase activity is defective in all these mutants. NADPH oxidase assembly is normal for P415H/P415L and T341K mutants where mutation occurs in the consensus sequences of NADPH- and FAD-binding sites, respectively. This is in accordance with their buried position in the three-dimensional model of the cytosolic Nox2 domain. FAD incorporation is abolished only in the T341K mutant explaining its absence of diaphorase activity. This demonstrates that NADPH oxidase assembly can occur without FAD incorporation. In addition, a defect of NADPH binding is a plausible explanation for the diaphorase activity inhibition in the P415H, P415L, and C537R mutants. In contrast, Cys-369, Gly-408, Leu-546, and Glu-568 are essential for NADPH oxidase complex assembly. However, according to their position in the three-dimensional model of the cytosolic domain of Nox2, only Cys-369 could be in direct contact with cytosolic factors during oxidase assembly. In addition, the defect in oxidase assembly observed in the C369R, G408E, G408R, and E568K mutants correlates with the lack of FAD incorporation. Thus, the NADPH oxidase assembly process and FAD incorporation are closely related events essential for the diaphorase activity of Nox2.
Figures




Similar articles
-
Role of putative second transmembrane region of Nox2 protein in the structural stability and electron transfer of the phagocytic NADPH oxidase.J Biol Chem. 2011 Aug 12;286(32):28357-69. doi: 10.1074/jbc.M111.220418. Epub 2011 Jun 9. J Biol Chem. 2011. PMID: 21659519 Free PMC article.
-
Crucial role of two potential cytosolic regions of Nox2, 191TSSTKTIRRS200 and 484DESQANHFAVHHDEEKD500, on NADPH oxidase activation.J Biol Chem. 2005 Apr 15;280(15):14962-73. doi: 10.1074/jbc.M500226200. Epub 2005 Jan 31. J Biol Chem. 2005. PMID: 15684431
-
Mutation at histidine 338 of gp91(phox) depletes FAD and affects expression of cytochrome b558 of the human NADPH oxidase.J Biol Chem. 1998 Oct 23;273(43):27879-86. doi: 10.1074/jbc.273.43.27879. J Biol Chem. 1998. PMID: 9774399
-
Hematologically important mutations: X-linked chronic granulomatous disease (third update).Blood Cells Mol Dis. 2010 Oct 15;45(3):246-65. doi: 10.1016/j.bcmd.2010.07.012. Epub 2010 Aug 21. Blood Cells Mol Dis. 2010. PMID: 20729109 Free PMC article. Review.
-
The molecular basis of chronic granulomatous disease.Springer Semin Immunopathol. 1998;19(4):417-34. doi: 10.1007/BF00792600. Springer Semin Immunopathol. 1998. PMID: 9618766 Review.
Cited by
-
Role of NADPH Oxidases in Stroke Recovery.Antioxidants (Basel). 2024 Aug 30;13(9):1065. doi: 10.3390/antiox13091065. Antioxidants (Basel). 2024. PMID: 39334724 Free PMC article. Review.
-
Antepenultimate residue at the C-terminus of NADPH oxidase RBOHD is critical for its function in the production of reactive oxygen species in Arabidopsis.J Zhejiang Univ Sci B. 2019 Sept.;20(9):713-727. doi: 10.1631/jzus.B1900105. J Zhejiang Univ Sci B. 2019. PMID: 31379142 Free PMC article.
-
Genetic disorders coupled to ROS deficiency.Redox Biol. 2015 Dec;6:135-156. doi: 10.1016/j.redox.2015.07.009. Epub 2015 Jul 17. Redox Biol. 2015. PMID: 26210446 Free PMC article. Review.
-
Molecular characterization of LC3-associated phagocytosis reveals distinct roles for Rubicon, NOX2 and autophagy proteins.Nat Cell Biol. 2015 Jul;17(7):893-906. doi: 10.1038/ncb3192. Epub 2015 Jun 22. Nat Cell Biol. 2015. Retraction in: Nat Cell Biol. 2024 Apr;26(4):660. doi: 10.1038/s41556-024-01383-1. PMID: 26098576 Free PMC article. Retracted.
-
The dehydrogenase region of the NADPH oxidase component Nox2 acts as a protein disulfide isomerase (PDI) resembling PDIA3 with a role in the binding of the activator protein p67 (phox.).Front Chem. 2015 Feb 4;3:3. doi: 10.3389/fchem.2015.00003. eCollection 2015. Front Chem. 2015. PMID: 25699251 Free PMC article.
References
-
- Balbridge C. W., Gerard R. W. (1933) Am. J. Physiol. 103, 235–236
-
- van den Berg J. M., van Koppen E., Ahlin A., Belohradsky B. H., Bernatowska E., Corbeel L., Español T., Fischer A., Kurenko-Deptuch M., Mouy R., Petropoulou T., Roesler J., Seger R., Stasia M. J., Valerius N. H., Weening R. S., Wolach B., Roos D., Kuijpers T. W. (2009) PLoS One 4, e5234. - PMC - PubMed
-
- Yu L., Zhen L., Dinauer M. C. (1997) J. Biol. Chem. 272, 27288–27294 - PubMed
-
- Aliverti A., Bruns C. M., Pandini V. E., Karplus P. A., Vanoni M. A., Curti B., Zanetti G. (1995) Biochemistry 34, 8371–8379 - PubMed
-
- Bedard K., Krause K. H. (2007) Physiol. Rev. 87, 245–313 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
