

ATP-dependent mechanism protects spectrin against glycation in human erythrocytes
- PMID: 20724481
- PMCID: PMC2962492
- DOI: 10.1074/jbc.M110.126896
ATP-dependent mechanism protects spectrin against glycation in human erythrocytes
Abstract
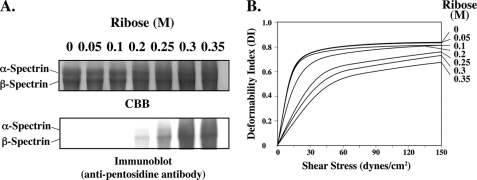
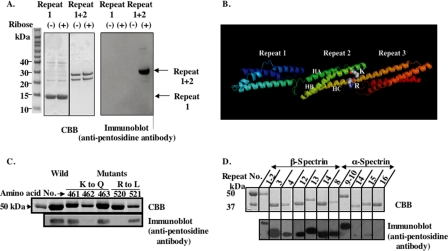
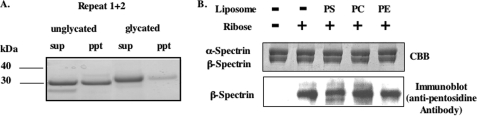
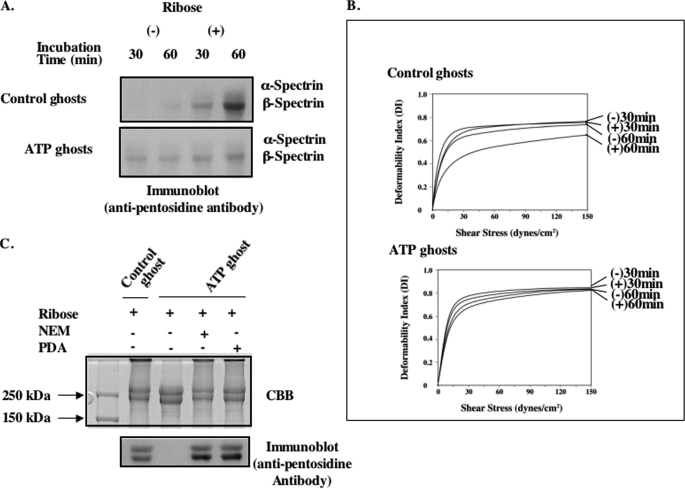
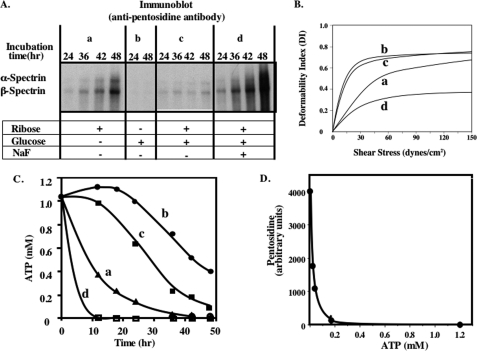
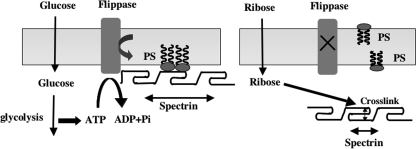
Human erythrocytes are continuously exposed to glucose, which reacts with the amino terminus of the β-chain of hemoglobin (Hb) to form glycated Hb, HbA1c, levels of which increase with the age of the circulating cell. In contrast to extensive insights into glycation of hemoglobin, little is known about glycation of erythrocyte membrane proteins. In the present study, we explored the conditions under which glucose and ribose can glycate spectrin, both on the intact membrane and in solution and the functional consequences of spectrin glycation. Although purified spectrin could be readily glycated, membrane-associated spectrin could be glycated only after ATP depletion and consequent translocation of phosphatidylserine (PS) from the inner to the outer lipid monolayer. Glycation of membrane-associated spectrin led to a marked decrease in membrane deformability. We further observed that only PS-binding spectrin repeats are glycated. We infer that the absence of glycation in situ is the consequence of the interaction of the target lysine and arginine residues with PS and thus is inaccessible for glycation. The reduced membrane deformability after glycation in the absence of ATP is likely the result of the inability of the glycated spectrin repeats to undergo the obligatory unfolding as a consequence of interhelix cross-links. We thus postulate that through the use of an ATP-driven phospholipid translocase (flippase), erythrocytes have evolved a protective mechanism against spectrin glycation and thus maintain their optimal membrane function during their long circulatory life span.
Figures

References
-
- Bierhaus A., Hofmann M. A., Ziegler R., Nawroth P. P. (1998) Cardiovasc. Res. 37, 586–600 - PubMed
-
- Sell D. R., Monnier V. M. (1989) J. Biol. Chem. 264, 21597–21602 - PubMed
-
- Grandhee S. K., Monnier V. M. (1991) J. Biol. Chem. 266, 11649–11653 - PubMed
-
- Bunn H. F., Gabbay K. H., Gallop P. M. (1978) Science 200, 21–27 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
