

Higher-order correlations in non-stationary parallel spike trains: statistical modeling and inference
- PMID: 20725510
- PMCID: PMC2906200
- DOI: 10.3389/fncom.2010.00016
Higher-order correlations in non-stationary parallel spike trains: statistical modeling and inference
Abstract
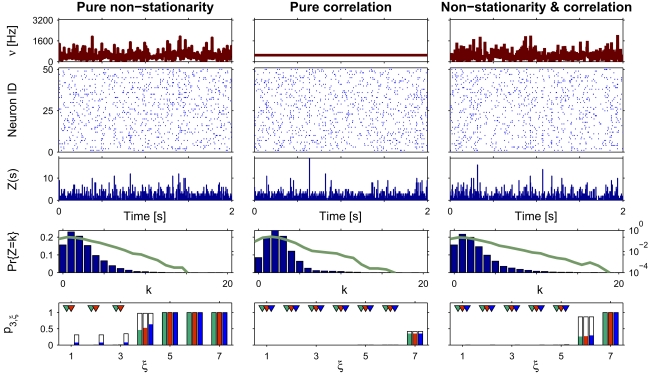
The extent to which groups of neurons exhibit higher-order correlations in their spiking activity is a controversial issue in current brain research. A major difficulty is that currently available tools for the analysis of massively parallel spike trains (N >10) for higher-order correlations typically require vast sample sizes. While multiple single-cell recordings become increasingly available, experimental approaches to investigate the role of higher-order correlations suffer from the limitations of available analysis techniques. We have recently presented a novel method for cumulant-based inference of higher-order correlations (CuBIC) that detects correlations of higher order even from relatively short data stretches of length T = 10-100 s. CuBIC employs the compound Poisson process (CPP) as a statistical model for the population spike counts, and assumes spike trains to be stationary in the analyzed data stretch. In the present study, we describe a non-stationary version of the CPP by decoupling the correlation structure from the spiking intensity of the population. This allows us to adapt CuBIC to time-varying firing rates. Numerical simulations reveal that the adaptation corrects for false positive inference of correlations in data with pure rate co-variation, while allowing for temporal variations of the firing rates has a surprisingly small effect on CuBICs sensitivity for correlations.
Keywords: higher-order correlations; multiple unit activity; non-stationarity; statistical population model.
Figures




Similar articles
-
A new method to infer higher-order spike correlations from membrane potentials.J Comput Neurosci. 2013 Oct;35(2):169-86. doi: 10.1007/s10827-013-0446-8. Epub 2013 Mar 10. J Comput Neurosci. 2013. PMID: 23474914 Free PMC article.
-
CuBIC: cumulant based inference of higher-order correlations in massively parallel spike trains.J Comput Neurosci. 2010 Aug;29(1-2):327-350. doi: 10.1007/s10827-009-0195-x. Epub 2009 Oct 28. J Comput Neurosci. 2010. PMID: 19862611 Free PMC article.
-
Modeling and analyzing higher-order correlations in non-Poissonian spike trains.J Neurosci Methods. 2012 Jun 30;208(1):18-33. doi: 10.1016/j.jneumeth.2012.04.015. Epub 2012 Apr 26. J Neurosci Methods. 2012. PMID: 22561088
-
Effect of stimulation on burst firing in cat primary auditory cortex.J Neurophysiol. 1995 Nov;74(5):1841-55. doi: 10.1152/jn.1995.74.5.1841. J Neurophysiol. 1995. PMID: 8592178
-
Data-driven significance estimation for precise spike correlation.J Neurophysiol. 2009 Mar;101(3):1126-40. doi: 10.1152/jn.00093.2008. Epub 2009 Jan 7. J Neurophysiol. 2009. PMID: 19129298 Free PMC article. Review.
Cited by
-
A new method to infer higher-order spike correlations from membrane potentials.J Comput Neurosci. 2013 Oct;35(2):169-86. doi: 10.1007/s10827-013-0446-8. Epub 2013 Mar 10. J Comput Neurosci. 2013. PMID: 23474914 Free PMC article.
-
Cell assemblies at multiple time scales with arbitrary lag constellations.Elife. 2017 Jan 11;6:e19428. doi: 10.7554/eLife.19428. Elife. 2017. PMID: 28074777 Free PMC article.
-
Exact analysis of the subthreshold variability for conductance-based neuronal models with synchronous synaptic inputs.ArXiv [Preprint]. 2023 Dec 28:arXiv:2304.09280v3. ArXiv. 2023. Update in: Phys Rev X. 2024 Jan-Mar;14(1):011021. doi: 10.1103/physrevx.14.011021. PMID: 37131877 Free PMC article. Updated. Preprint.
-
ASSET: Analysis of Sequences of Synchronous Events in Massively Parallel Spike Trains.PLoS Comput Biol. 2016 Jul 15;12(7):e1004939. doi: 10.1371/journal.pcbi.1004939. eCollection 2016 Jul. PLoS Comput Biol. 2016. PMID: 27420734 Free PMC article.
-
LARGE-SCALE MULTIPLE INFERENCE OF COLLECTIVE DEPENDENCE WITH APPLICATIONS TO PROTEIN FUNCTION.Ann Appl Stat. 2021 Jun;15(2):902-924. doi: 10.1214/20-aoas1431. Epub 2021 Jul 12. Ann Appl Stat. 2021. PMID: 35910493 Free PMC article.
References
-
- Abeles M. (1991). Corticonics: Neural Circuits of the Cerebral Cortex, 1st Edn.Cambridge: Cambridge University Press
-
- Aertsen A., Gerstein G., Habib M., Palm G. (1989). Dynamics of neuronal firing correlation: modulation of “effective connectivity”. J. Neurophysiol. 61, 900–917 - PubMed
LinkOut - more resources
Full Text Sources