

Review
doi: 10.1007/s00018-010-0482-8.
Epub 2010 Aug 21.
Crustacean neuropeptides
Affiliations
- PMID: 20725764
- PMCID: PMC11115526
- DOI: 10.1007/s00018-010-0482-8
Item in Clipboard
Review
Crustacean neuropeptides
Cell Mol Life Sci.
2010 Dec.
Abstract
Crustaceans have long been used for peptide research. For example, the process of neurosecretion was first formally demonstrated in the crustacean X-organ-sinus gland system, and the first fully characterized invertebrate neuropeptide was from a shrimp. Moreover, the crustacean stomatogastric and cardiac nervous systems have long served as models for understanding the general principles governing neural circuit functioning, including modulation by peptides. Here, we review the basic biology of crustacean neuropeptides, discuss methodologies currently driving their discovery, provide an overview of the known families, and summarize recent data on their control of physiology and behavior.
Figures

Nucleotide and deduced amino acid sequences of Homarus americanus (Homam) prepro-sulfakinin. A1 Nucleotide sequence of Homam-prepro-sulfakinin cDNA (accession no. EF418605). The open reading frame of the cDNA, including the stop codon, is shown in black font, with two 3′ polyadenylation signal sequences indicated by underline in black. A2 Deduced amino acid sequence of Homam-prepro-sulfakinin. The signal peptide is shown in grey, with prohormone convertase cleavage loci shown in black. The two encoded sulfakinin isoforms are shown in red, with additional precursor-related peptides shown in blue. The asterisk indicates the position of the stop codon. B Putative processing scheme resulting in the production of the two isoforms of Homam-sulfakinin from its precursor protein. The mature conformations of the two sulfakinin isoforms (Homam-SK I and II) are colored red. Figure modified from Dickinson et al. [158]
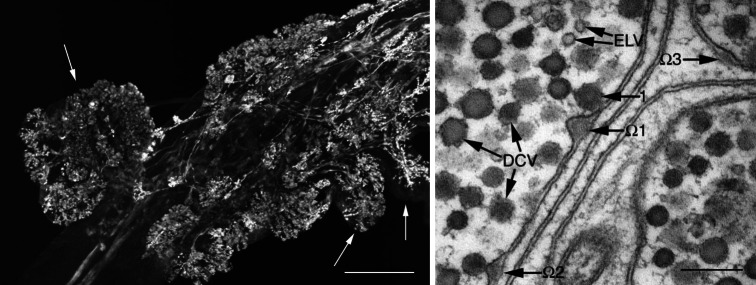
General organization of a crustacean neuroendocrine organ; the anterior cardiac plexus (ACP) of Cancer productus is used as an example. (Left panel) The Cancer productus ACP is composed of nerve terminals contained within blister-like protuberances of the anterior cardiac nerve sheath; these are in direct contact with the circulatory system. The confocal image shown illustrates the presence of FMRFamide-like immunoreactivity within these protuberances, several prominent ones indicated by arrows. Scale bar 100 µm. (Right panel) Transmission electron micrograph illustrating morphological correlates of hormone secretion in the Cancer productus ACP. Both dense-core vesicles (DCV), which are likely peptidergic, and electron-lucent vesicles (ELV) are present in these terminals. In this image, one DCV (1) is docked to the plasma membrane, while several others have fused with the membrane and are in the process of exocytosing their contents, creating characteristic ultrastuctural features, i.e., omega (Ω)-figures, on the plasma membrane (Ω2, Ω3). The docked DCV and the three Ω-figures visible in this micrograph create a pseudo-time course of peptide hormone secretion. A DCV first docks to the plasma membrane (1), then fuses with it, releasing its dense-core and forming an Ω-figure (Ω1). The membrane of the DCV is rapidly incorporated into the plasma membrane of the terminal, and the Ω-figure subsides (Ω2 and Ω3). Scale bar 200 nm. Figure modified from Christie et al. [8]

Direct tissue MALDI-FTMS used for neuropeptide profiling. In this example, a single stomatogastric ganglion (STG) from an a embryonic or b adult Homarus americanus was analyzed. The STG, which is small enough to be analyzed as a whole tissue, was freshly dissected, rinsed in acidified methanol, desalted, and co-crystallized with 2,5-dihydroxybenzoic acid as the MALDI matrix. Figure modified from Cape et al. [46]; used with permission

a MS/MS spectrum of [Ala13]-orcokinin from the brain of Cancer borealis, measured using a MALDI-TOF/TOF instrument. MS/MS was carried out using air as the collision gas and a 2-kV collision energy. b MS/MS spectrum of an [Ala13]-orcokinin standard, measured using a MALDI-FTMS instrument. MS/MS was carried out using SORI-CID with argon as the collision gas and a Vp amplitude of 6.5 V. The lower energy SORI-CID conditions yield spectra dominated by Asp-Xxx cleavages (cleavages C-terminal to aspartate residues) and fewer product ions that can be used for peptide sequencing. Figure 4a modified from Chen et al. [47]; used with permission

Modulation of the heartbeat in Homarus americanus by TNRNFLRFamide involves complex effects at multiple sites. a, b Schematic diagrams of the preparations used to record muscle contraction and activity of the cardiac ganglion. a Contraction of an isolated whole heart was monitored with a force transducer; motor output of the cardiac ganglion was recorded simultaneously with a suction electrode on a motor nerve. b In other preparations, the cardiac ganglion was removed from the heart, and motor output was recorded on the motor nerves in the isolated cardiac ganglion. c Perfusion of TNRNFLRFamide through the heart increases both amplitude and frequency of heart contractions. C1 and C2 are higher speed recordings from the regions shown in c, illustrating the increases in both amplitude and frequency recorded during TNRNFLRFamide perfusion (yellow bar in c; C2). d The effect of TNRNFLRFamide on contraction amplitude in the whole heart is dose-dependent, with significant effects at concentrations as low as 10−10 M. e Cycle frequency of the motor bursts recorded in the whole heart (blue bars) increases in response to perfusion with TNRNFLRFamide, with a threshold of ~10−10 M. In contrast, superfusion of the same peptide over the isolated CG causes a decrease in cycle frequency (green bars). Moreover, effects of this peptide on the isolated CG are seen only at much higher concentrations (e.g., 10−8 M) than those on the whole heart. Scale bars 1.0 g, 100 s in c, 2.5 s in C1 and C2. Figure modified from Stevens et al. [273]
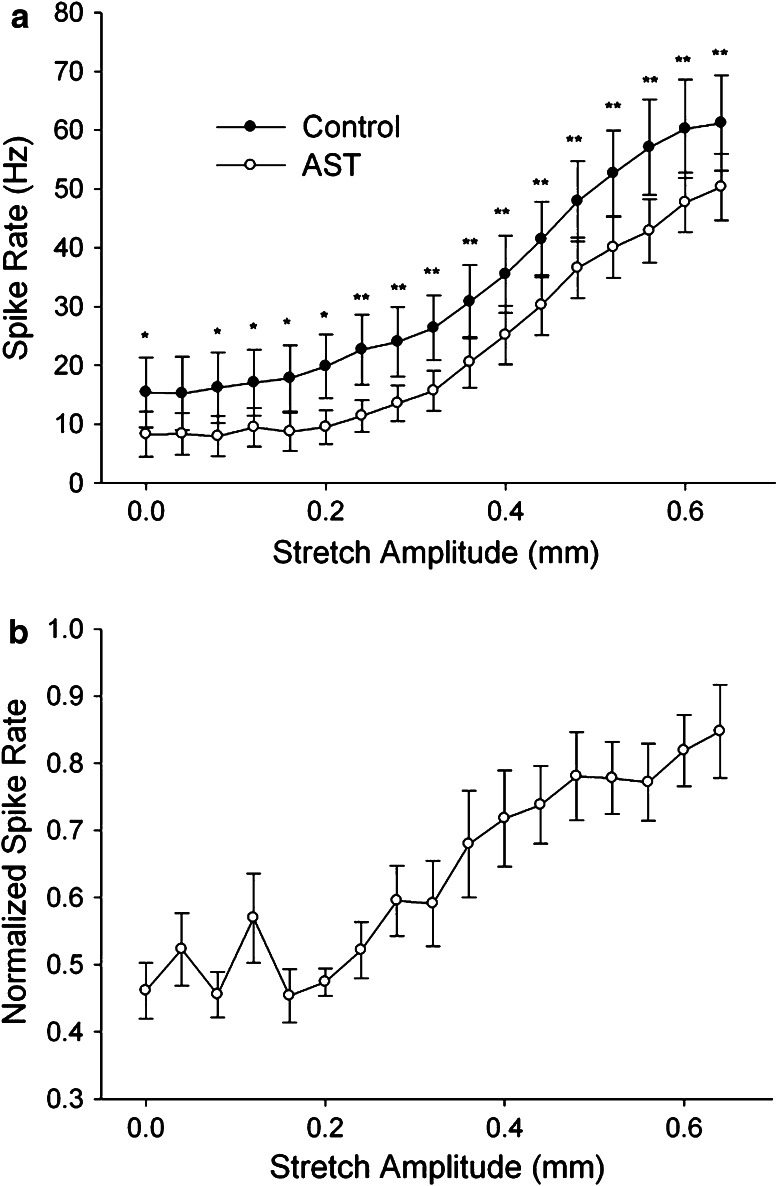
A-type allatostatin (A-AST) decreases the responses of the gastro-pyloric stretch receptor (GPR) in the Cancer borealis stomatogastric system. a Spike frequency in response to stretch increases as a function of stretch amplitude, but is lower at all amplitudes in the presence of AST. b Spike frequency in A-AST is shown normalized to the response in control saline at each stretch amplitude, showing that the effects of A-AST are greater at larger stretch amplitudes. Figure modified from Birmingham et al. [297]; used with permission
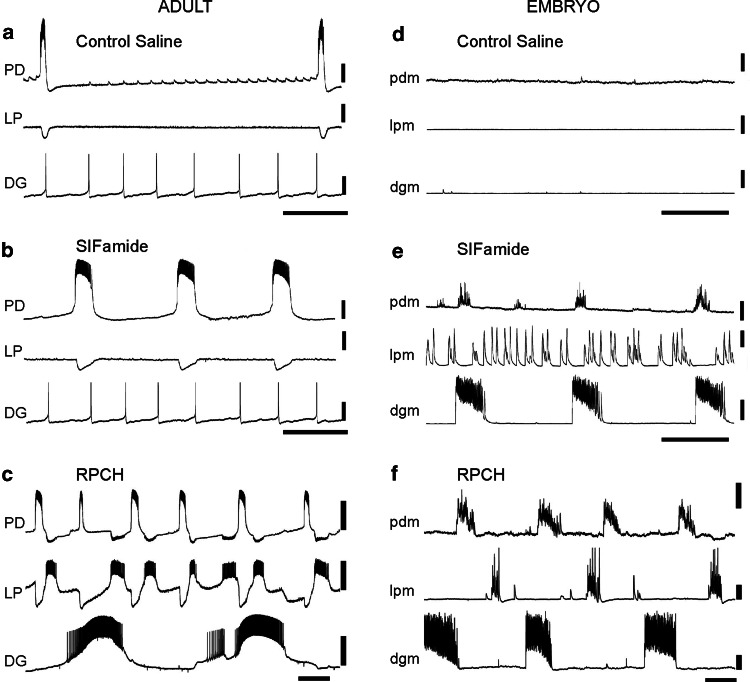
Peptides modulate the same system differently in adults (a, b, c) and embryos (d, e, f) of Homarus americanus. (a) When inputs from the anterior ganglia are blocked in the adult, the only rhythmically active neurons in the STG are the PD neurons, which fire in regular bursts at a low cycle frequency. b Superfusion of the isolated STG with SIFamide activates bursting in the PD neurons, but has little effect on the other two neurons shown, the LP neuron of the pyloric network, and the DG neuron from the gastric mill network. c RPCH causes an increase in activity in all three neurons shown. d In the embryo, recordings are from muscles innervated by the same neurons whose activity is shown in the adult neuronal recordings. In control saline, when modulatory inputs are blocked, none of the neurons are rhythmically active. e Superfusion with SIFamide activates all three neurons, with the PD and DG innervated muscles (pdm, lpm) firing in bursts that are more or less synchronous, while the LP-innervated muscle (dgm) is activated more or less tonically. This contrasts with the adult, in which only one of the neurons (i.e., PD) is activated. f RPCH activates activity in all three muscles; however, in contrast to what is seen in the adult, they all fire with the same cycle frequency. c and f are from different preparations than a/b and d/e, respectively; note also that they are on slightly different time scales. Scale bars 5 mV, 5 s. Figure modified from Rehm et al. [228]; used with permission
References
-
- Christie AE, Skiebe P, Marder E. Matrix of neuromodulators in neurosecretory structures of the crab Cancer borealis . J Exp Biol. 1995;198:2431–2439. - PubMed
-
- Hooper SL, DiCaprio RA. Crustacean motor pattern generator networks. Neurosignals. 2004;13:50–69. - PubMed
-
- Mulloney B, Skinner FK, Namba H, Hall WM. Intersegmental coordination of swimmeret movements: mathematical models and neural circuits. Ann NY Acad Sci. 1998;860:266–280. - PubMed
-
- Cooke IM. Reliable, responsive pacemaking and pattern generation with minimal cell numbers: the crustacean cardiac ganglion. Biol Bull. 2002;202:108–136. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
