

Review
doi: 10.1111/j.1349-7006.2010.01683.x.
The microRNA pathway and cancer
Affiliations
- PMID: 20726859
- PMCID: PMC11159795
- DOI: 10.1111/j.1349-7006.2010.01683.x
Item in Clipboard
Review
The microRNA pathway and cancer
Cancer Sci.
2010 Nov.
Abstract
MicroRNAs (miRNAs) are ∼22nt long, non-coding RNAs that guide post-transcriptional gene silencing of their target genes and regulate diverse biological processes including cancer. miRNAs do not act alone, but require assembly into RNA-induced silencing complex (RISC). In this review, we summarize how miRNAs are produced, assembled into RISC, and regulate target mRNAs, and discuss how the miRNA pathway is involved in cancer.
© 2010 Japanese Cancer Association.
Figures
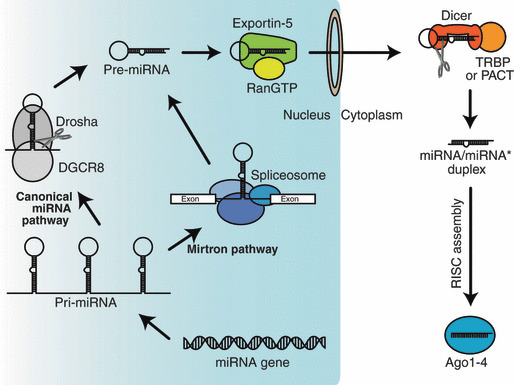
Biogenesis of microRNA (miRNA). miRNAs are encoded in the genome and upon transcription give rise to primary miRNAS (pri‐miRNAs). The Drosha/DiGeorge syndrome critical region 8 protein (DGCR8) complex or the spliceosome catalyzes pri‐ to precursor miRNA (pre‐miRNA) conversion. Exportin‐5/RanGTP complex exports pre‐miRNAs from the nucleus to the cytoplasm. Dicer interacts with TAR RNA‐binding protein (TRBP) or PKR activating protein (PACT) and cleaves pre‐miRNAs into miRNA/miRNA* duplexes. These duplexes are assembled into Argonaute (Ago)1–4, and eventually form mature RNA‐induced silencing complexes (RISCs), which contain only single‐stranded guides.
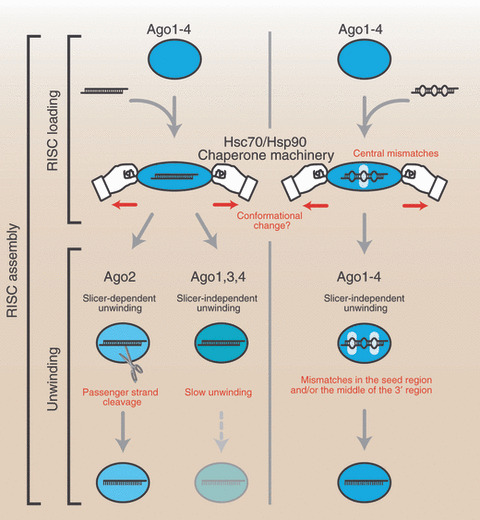
RNA‐induced silencing complex (RISC) assembly can be divided into at least two steps, RISC loading and unwinding. The former prefers central mismatches in microRNA (miRNA)/miRNA* duplexes. Perfectly complementary siRNA duplexes can also be loaded into Argonaute (Ago)1–4, but duplexes without central mismatches are not favored for RISC loading. The RISC loading of small RNA duplexes is ATP‐dependent and facilitated by Hsc70/Hsp90 chaperone machinery. Mismatches in the seed region and/or the middle of the 3′ region enhance the slicer‐independent unwinding efficiency of all four Ago proteins. In contrast, only Ago2 can efficiently unwind perfectly complementary duplexes by passenger strand cleavage.
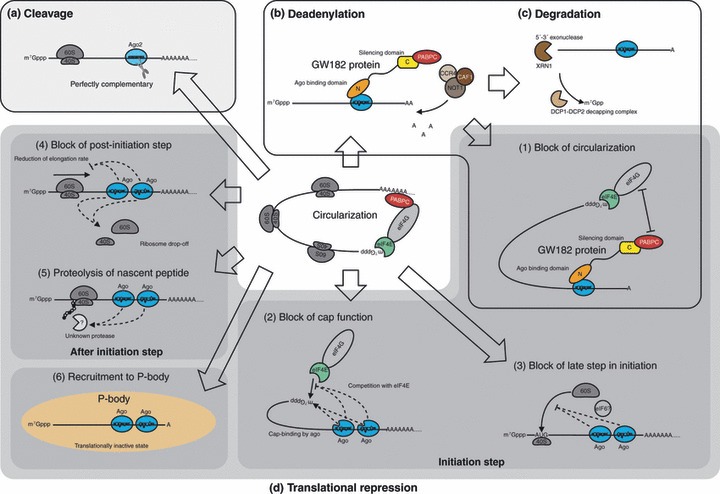
RNA‐induced silencing (RISC) regulates gene expression through various mechanisms: (a) cleavage; (b) deadenylation; (c) degradation; and (d) translational repression of target mRNA. (1–6) Proposed mechanisms of translational repression. Mechanisms 1–3 target the initiation step of translation; mechanisms 4 and 5 target the steps after initiation. AAA..., poly(A) tail; Ago, Argonaute; C, C‐terminal domain of GW182 protein: silencing domain; CAF1, CCR4 associated factor 1; CCR4, carbon catabolite repression 4; N, N‐terminal domain of GW182 protein: Ago binding domain; NOT1, negative on TATA‐less 1; PABPC, cytoplasmic poly(A) binding protein; P‐body, processing body; XRN1, 5′‐exoribonuclease 1.

MicroRNA (miRNA) pathway and cancer. (a) Genomic abnormalities, such as chromosomal translocations and point mutations, can attenuate or stimulate miRNA transcription leading to an increase or decrease of primary miRNA (pri‐miRNA). (b) Mutations in a miRNA gene can lead to RNA‐induced silencing (RISC) assembly abnormalities. For example, when nucleotides that establish the thermodynamic asymmetry are substituted, this can lead to “flipped” strand selection, resulting in the miRNA* strand, instead of the miRNA strand, being favored for RISC incorporation. (c) Furthermore, single nucleotide polymorphisms or mutations in either the miRNA or the target RNA can abrogate proper target recognition, especially when located within the seed region. Consequently, genes might escape being regulated by RISCs or perhaps even become differently regulated as a result of erroneous targeting. (d) Genomic abnormalities can also result in aberrant miRNA processing. Some pri‐miRNAs require additional proteins for efficient conversion. p53 mutants affect the interaction between p68/p72 RNA helicases and Drosha and this decreases pri‐ to precursor miRNA (pre‐miRNA) conversion of a subset of miRNAs. Additionally, p68 associates with receptor‐regulated Smads. This interaction has been shown to be important for efficient pri‐ to pre‐miRNA conversion of another subset of miRNAs. In the cytoplasm, TAR RNA‐binding protein (TRBP) is phosphorylated by MAPK/Erk signaling. The phosphorylation of TRBP increases pre‐miRNA to miRNA/miRNA* conversion for general miRNAs, but it decreases conversion for let‐7 family miRNAs. AAA..., poly(A) tail; Ago, Argonaute; DGCR8, DiGeorge syndrome critical region 8 protein.
Similar articles
-
Intracellular and extracellular microRNA: An update on localization and biological role.Prog Histochem Cytochem. 2016 Nov;51(3-4):33-49. doi: 10.1016/j.proghi.2016.06.001. Epub 2016 Jun 25. Prog Histochem Cytochem. 2016. PMID: 27396686 Review.
-
Nuclear functions of mammalian MicroRNAs in gene regulation, immunity and cancer.Mol Cancer. 2018 Feb 22;17(1):64. doi: 10.1186/s12943-018-0765-5. Mol Cancer. 2018. PMID: 29471827 Free PMC article. Review.
-
MicroRNA Expression: Protein Participants in MicroRNA Regulation.Methods Mol Biol. 2017;1617:27-37. doi: 10.1007/978-1-4939-7046-9_2. Methods Mol Biol. 2017. PMID: 28540674 Review.
-
MicroRNAs: molecular features and role in cancer.Front Biosci (Landmark Ed). 2012 Jun 1;17(7):2508-40. doi: 10.2741/4068. Front Biosci (Landmark Ed). 2012. PMID: 22652795 Free PMC article. Review.
-
Target binding triggers hierarchical phosphorylation of human Argonaute-2 to promote target release.Elife. 2022 May 31;11:e76908. doi: 10.7554/eLife.76908. Elife. 2022. PMID: 35638597 Free PMC article.
Cited by
-
MicroRNA-regulated protein-protein interaction networks and their functions in breast cancer.Int J Mol Sci. 2013 May 30;14(6):11560-606. doi: 10.3390/ijms140611560. Int J Mol Sci. 2013. PMID: 23722663 Free PMC article.
-
Expression and clinical significance of microRNA-1246 in human oral squamous cell carcinoma.Med Sci Monit. 2015 Mar 16;21:776-81. doi: 10.12659/MSM.892508. Med Sci Monit. 2015. PMID: 25791131 Free PMC article.
-
The Expanding Role of Cancer Stem Cell Marker ALDH1A3 in Cancer and Beyond.Cancers (Basel). 2023 Jan 13;15(2):492. doi: 10.3390/cancers15020492. Cancers (Basel). 2023. PMID: 36672441 Free PMC article. Review.
-
Comprehensive Review of Genetic Association Studies and Meta-Analysis on polymorphisms in microRNAs and Urological Neoplasms Risk.Sci Rep. 2018 Feb 28;8(1):3776. doi: 10.1038/s41598-018-21749-4. Sci Rep. 2018. PMID: 29491365 Free PMC article. Review.
-
The N domain of Argonaute drives duplex unwinding during RISC assembly.Nat Struct Mol Biol. 2012 Jan 10;19(2):145-51. doi: 10.1038/nsmb.2232. Nat Struct Mol Biol. 2012. PMID: 22233755
References
-
- Borchert GM, Lanier W, Davidson BL. RNA polymerase III transcribes human microRNAs. Nat Struct Mol Biol 2006; 13: 1097–101. - PubMed
-
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004; 116: 281–97. - PubMed
-
- Lee Y, Ahn C, Han J et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003; 425: 415–9. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
