

New insights into the enigma of immunoglobulin D
- PMID: 20727035
- PMCID: PMC3048779
- DOI: 10.1111/j.1600-065X.2010.00929.x
New insights into the enigma of immunoglobulin D
Abstract
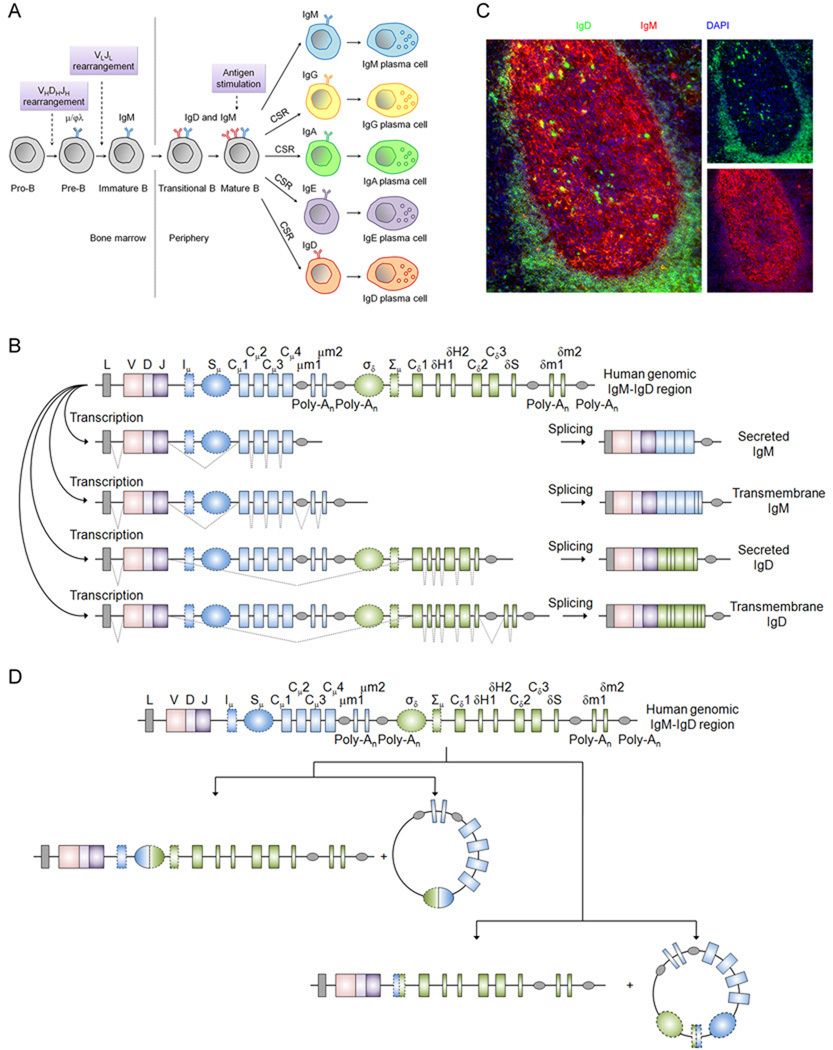
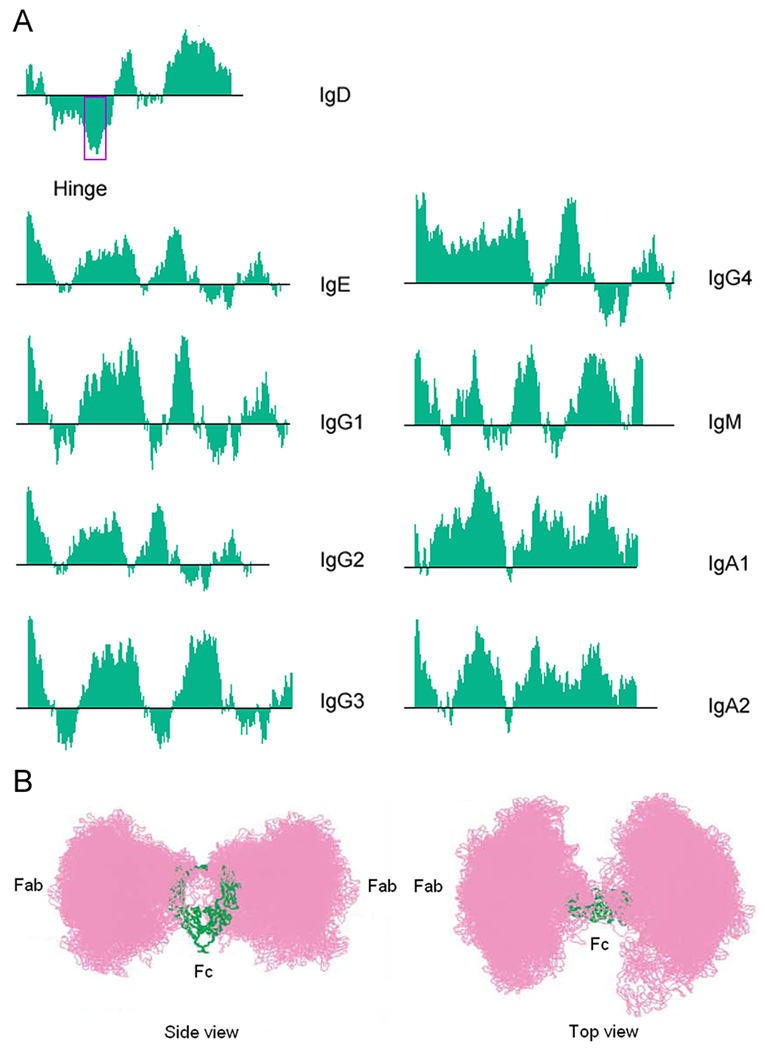
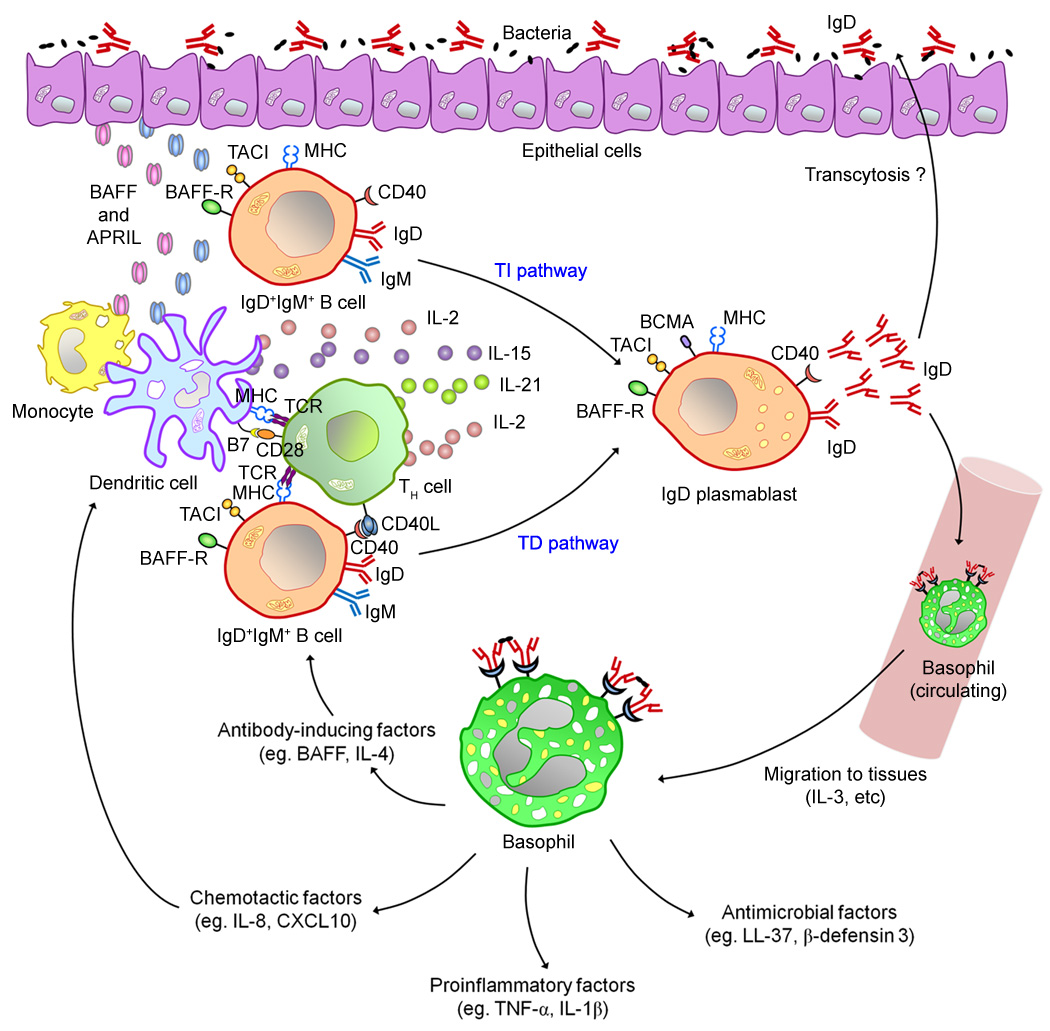
Immunoglobulin D (IgD) has remained a mysterious antibody class for almost half a century. IgD was initially thought to be a recently evolved Ig isotype expressed only by some mammalian species, but recent discoveries in fishes and amphibians demonstrate that IgD was present in the ancestor of all jawed vertebrates and has important immunological functions. The structure of IgD has been very dynamic throughout evolution. Mammals can express IgD through alternative splicing and class switch recombination. Active cell-dependent and T-cell-independent IgM-to-IgD class switching takes place in a unique subset of human B cells from the upper aerodigestive mucosa, which provides a layer of mucosal protection by interacting with many pathogens and their virulence factors. Circulating IgD can bind to myeloid cells such as basophils and induce antimicrobial, inflammatory, and B-cell-stimulating factors upon cross-linking, which contributes to not only immune surveillance but also inflammation and tissue damage when this pathway is overactivated under pathological conditions. Recent research shows that IgD is an important immunomodulator that orchestrates an ancestral surveillance system at the interface between immunity and inflammation.
Figures

References
-
- Black CA. A brief history of the discovery of the immunoglobulins and the origin of the modern immunoglobulin nomenclature. Immunol Cell Biol. 1997;75:65–68. - PubMed
-
- Finkelman FD, van Boxel JA, Asofsky R, Paul WE. Cell membrane IgD: demonstration of IgD on human lymphocytes by enzyme-catalyzed iodination and comparison with cell surface Ig of mouse, guinea pig, and rabbit. J Immunol. 1976;116:1173–1181. - PubMed
-
- Martin LN, Leslie GA, Hindes R. Lymphocyte surface IgD and IgM in non-human primates. Int Arch Allergy Appl Immunol. 1976;51:320–329. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
