

Multifactorial determinants of protein expression in prokaryotic open reading frames
- PMID: 20727358
- PMCID: PMC2963087
- DOI: 10.1016/j.jmb.2010.08.010
Multifactorial determinants of protein expression in prokaryotic open reading frames
Abstract
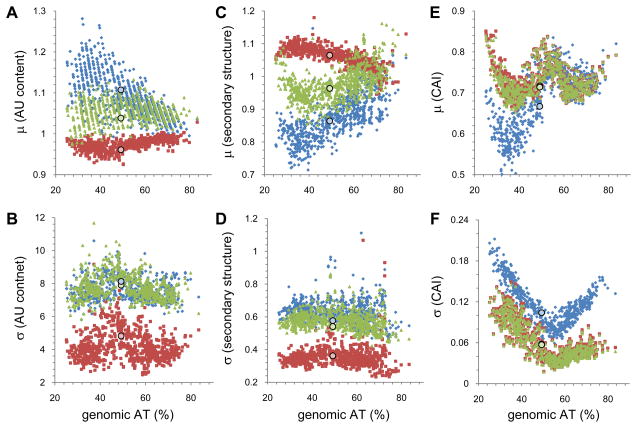
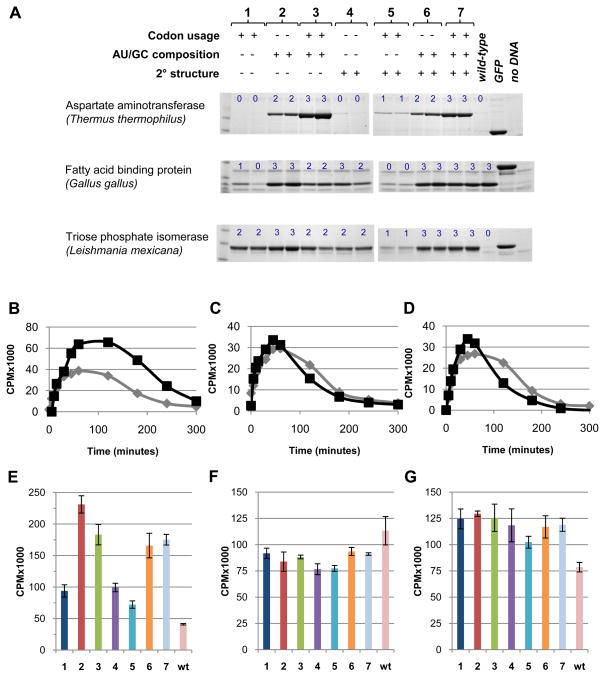
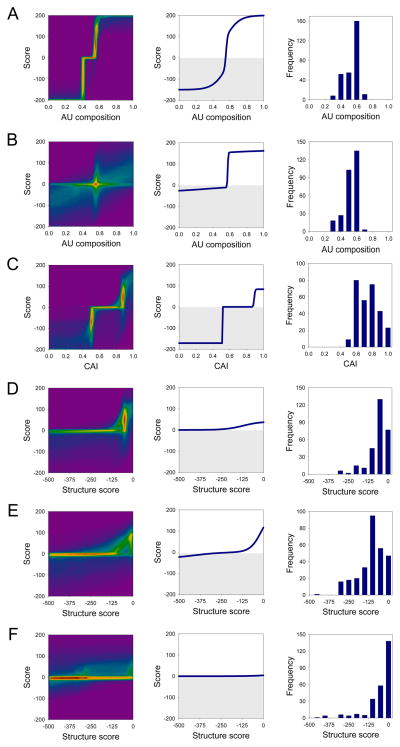
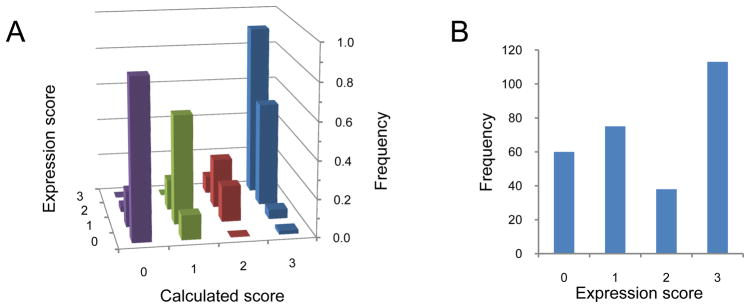
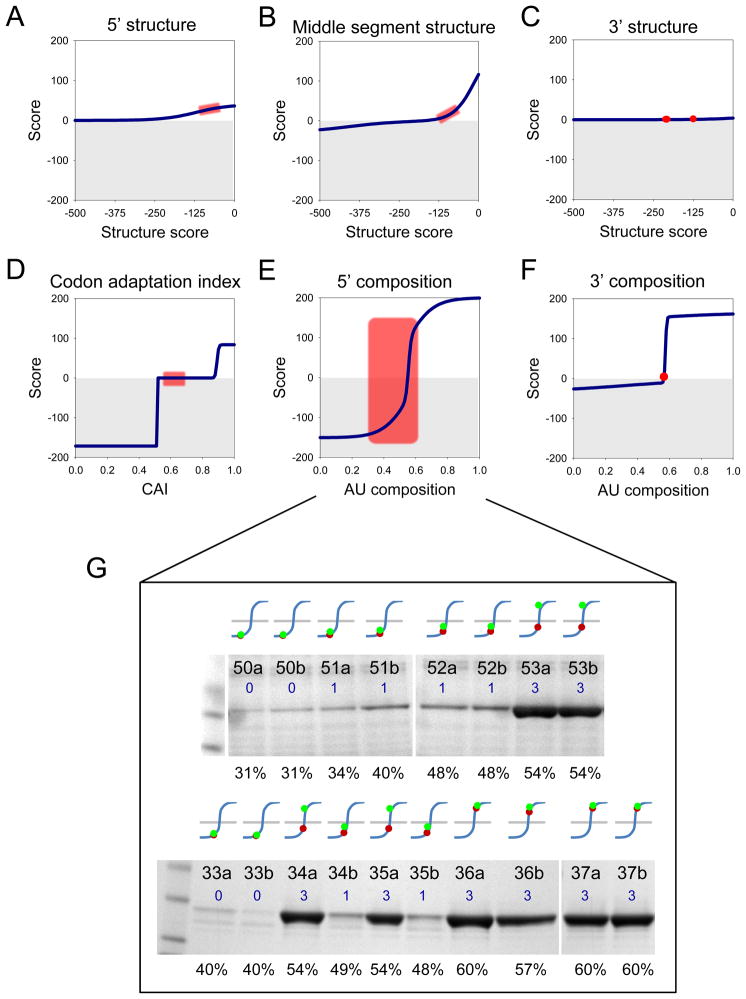
A quantitative description of the relationship between protein expression levels and open reading frame (ORF) nucleotide sequences is important for understanding natural systems, designing synthetic systems, and optimizing heterologous expression. Codon identity, mRNA secondary structure, and nucleotide composition within ORFs markedly influence expression levels. Bioinformatic analysis of ORF sequences in 816 bacterial genomes revealed that these features show distinct regional trends. To investigate their effects on protein expression, we designed 285 synthetic genes and determined corresponding expression levels in vitro using Escherichia coli extracts. We developed a mathematical function, parameterized using this synthetic gene data set, which enables computation of protein expression levels from ORF nucleotide sequences. In addition to its practical application in the design of heterologous expression systems, this equation provides mechanistic insight into the factors that control translation efficiency. We found that expression is strongly dependent on the presence of high AU content and low secondary structure in the ORF 5' region. Choice of high-frequency codons contributes to a lesser extent. The 3' terminal AU content makes modest, but detectable contributions. We present a model for the effect of these factors on the three phases of ribosomal function: initiation, elongation, and termination.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest
The authors declare that they have no conflict of interest.
Figures
References
-
- Carothers JM, Goler JA, Keasling JD. Chemical synthesis using synthetic biology. Curr Opin Biotechnol. 2009;20:498–503. - PubMed
-
- Jana S, Deb JK. Strategies for efficient production of heterologous proteins in Escherichia coli. Appl Microbiol Biotechnol. 2005;67:289–298. - PubMed
-
- Winkler WC, Breaker RR. Regulation of bacterial gene expression by riboswitches. Annu Rev Microbiol. 2005;59:487–517. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
