

Differential requirement of lipid rafts for FcγRIIA mediated effector activities
- PMID: 20728077
- PMCID: PMC2975250
- DOI: 10.1016/j.cellimm.2010.07.011
Differential requirement of lipid rafts for FcγRIIA mediated effector activities
Abstract
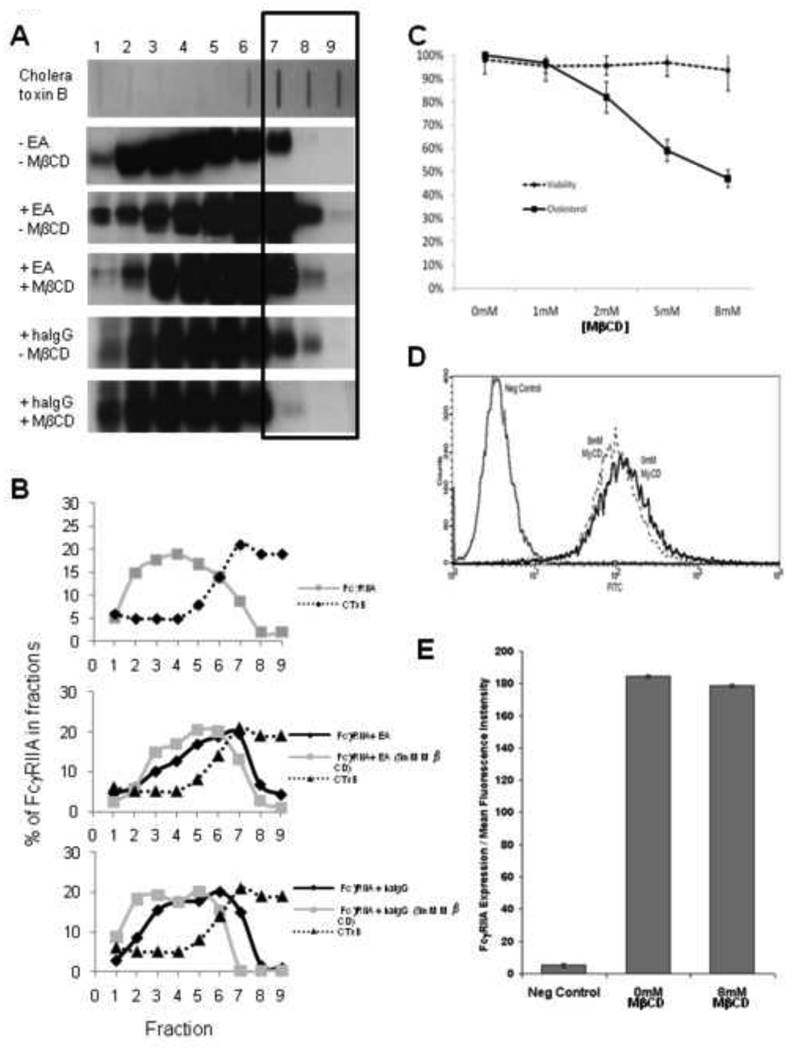
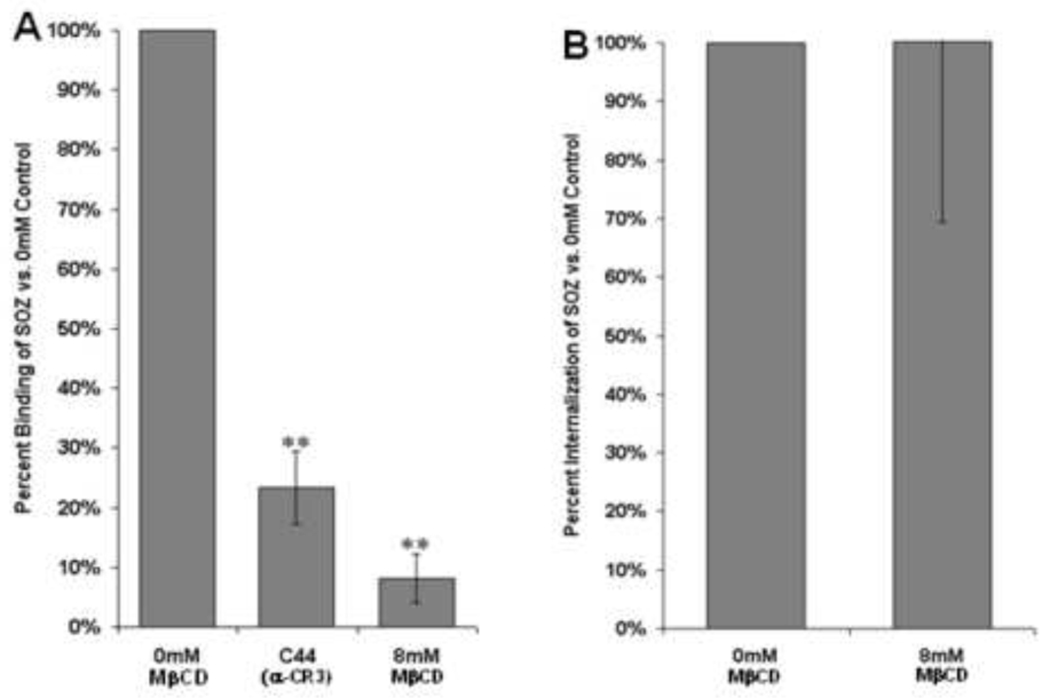
Immunoglobulin G (IgG) dependent activities are important in host defense and autoimmune diseases. Various cell types including macrophages and neutrophils contribute to pathogen destruction and tissue damage through binding of IgG to Fcγ receptors (FcγR). One member of this family, FcγRIIA, is a transmembrane glycoprotein known to mediate binding and internalization of IgG-containing targets. FcγRIIA has been observed to translocate into lipids rafts upon binding IgG-containing targets. We hypothesize that lipid rafts participate to different extents in binding and internalizing targets of different sizes. We demonstrate that disruption of lipid rafts with 8mM methyl-β-cyclodextrin (MβCD) nearly abolishes binding (91% reduction) and phagocytosis (60% reduction) of large IgG-coated targets. Conversely, binding and internalization of small IgG-complexes is less dependent on lipid rafts (49% and 17% inhibition at 8mM MβCD, respectively). These observations suggest that differences between phagocytosis and endocytosis may arise as early as the initial stages of ligand recognition.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures



References
-
- Gavin AL, Barnes N, Dijstelbloem HM, Hogarth PM. Identification of the mouse IgG3 receptor: implications for antibody effector function at the interface between innate and adaptive immunity. J Immunol. 1998;160:20–23. - PubMed
-
- Joshi T, Butchar JP, Tridandapani S. Fcgamma receptor signaling in phagocytes. Int J Hematol. 2006;84:210–216. - PubMed
-
- Hulett MD, Hogarth PM. Molecular basis of Fc receptor function. Adv Immunol. 1994;57:1–127. - PubMed
-
- Ravetch JV, Kinet JP. Fc receptors. Annu Rev Immunol. 1991;9:457–492. - PubMed
-
- Ganesan LP, Joshi T, Fang H, Kutala VK, Roda J, Trotta R, Lehman A, Kuppusamy P, Byrd JC, Carson WE, Caligiuri MA, Tridandapani S. FcgammaR-induced production of superoxide and inflammatory cytokines is differentially regulated by SHIP through its influence on PI3K and/or Ras/Erk pathways. Blood. 2006;108:718–725. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
