

Transmission of plant-pathogenic bacteria by nonhost seeds without induction of an associated defense reaction at emergence
- PMID: 20729326
- PMCID: PMC2953029
- DOI: 10.1128/AEM.01098-10
Transmission of plant-pathogenic bacteria by nonhost seeds without induction of an associated defense reaction at emergence
Abstract
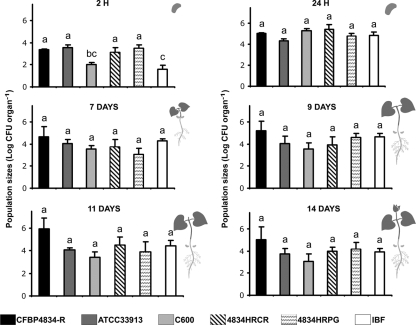
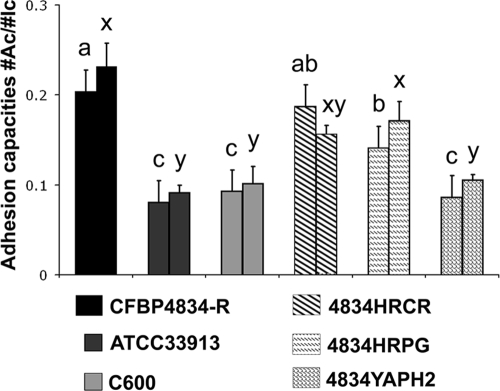
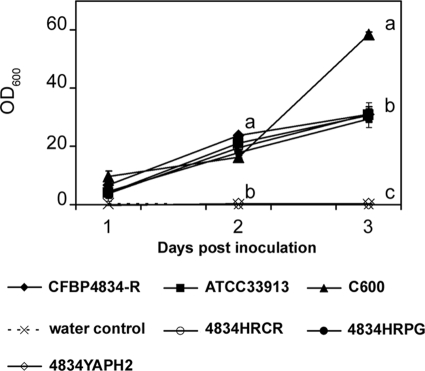
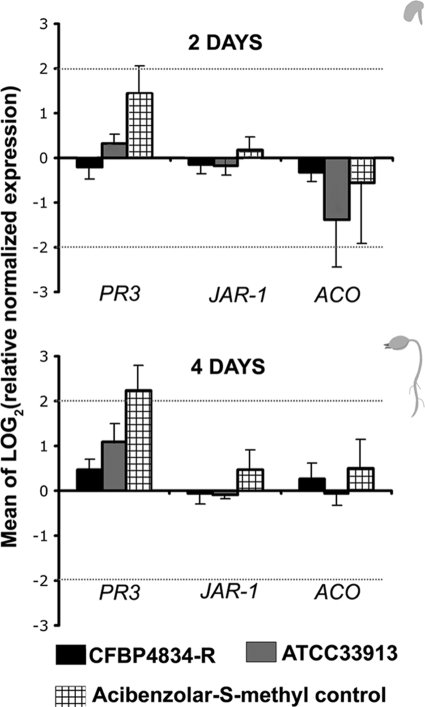
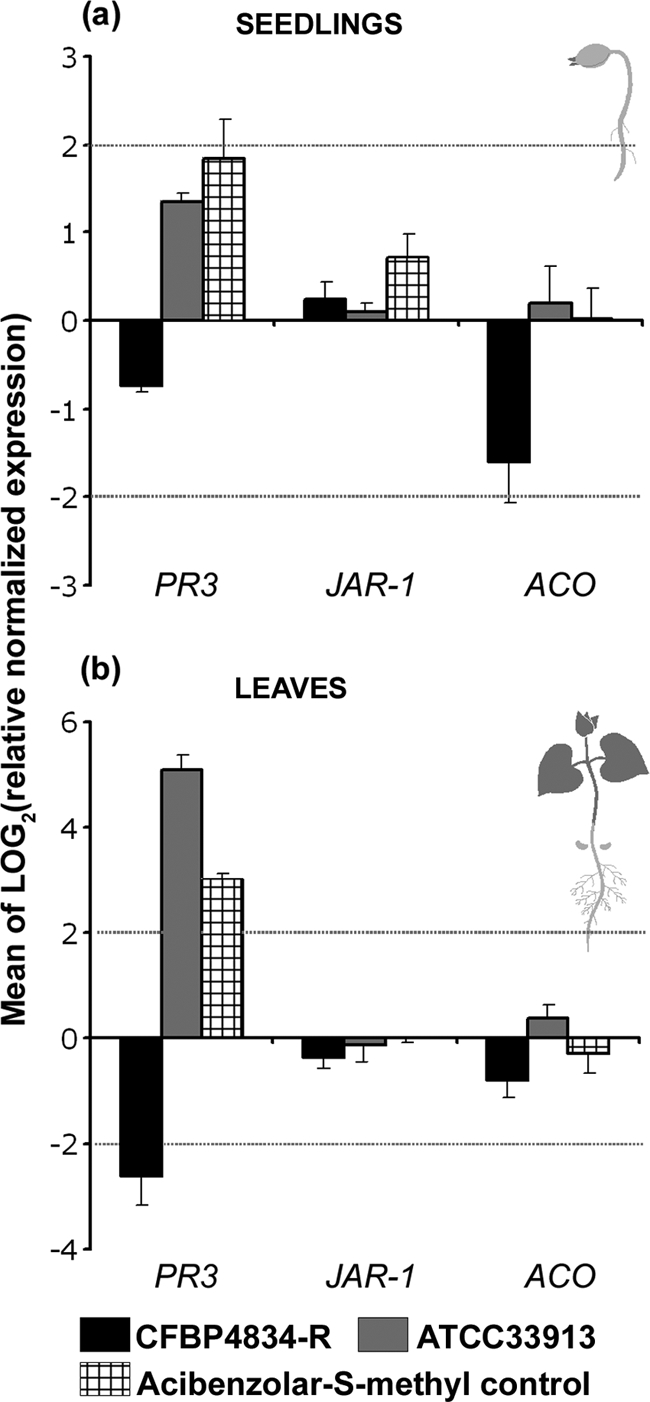
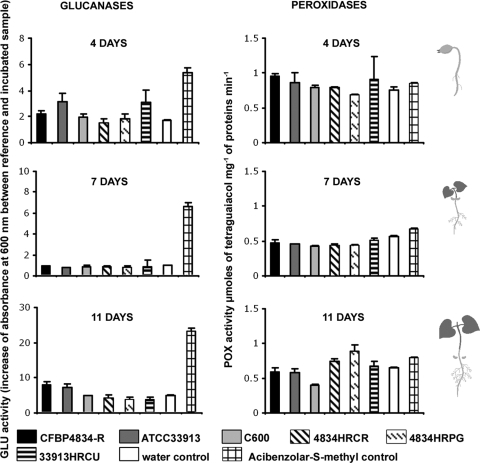
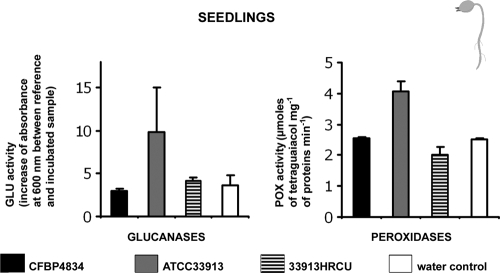
An understanding of the mechanisms involved in the different steps of bacterial disease epidemiology is essential to develop new control strategies. Seeds are the passive carriers of a diversified microbial cohort likely to affect seedling physiology. Among seed-borne plant-pathogenic bacteria, seed carriage in compatible situations is well evidenced. The aims of our work are to determine the efficiency of pathogen transmission to seeds of a nonhost plant and to evaluate bacterial and plant behaviors at emergence. Bacterial transmission from flowers to seeds and from seeds to seedlings was measured for Xanthomonas campestris pv. campestris in incompatible interactions with bean. Transmissions from seeds to seedlings were compared for X. campestris pv. campestris, for Xanthomonas citri pv. phaseoli var. fuscans in compatible interactions with bean, and for Escherichia coli, a human pathogen, in null interactions with bean. The induction of defense responses was monitored by using reverse transcription and quantitative PCR (RT-qPCR) of genes representing the main signaling pathways and assaying defense-related enzymatic activities. Flower inoculations resulted in a high level of bean seed contamination by X. campestris pv. campestris, which transmitted efficiently to seedlings. Whatever the type of interaction tested, dynamics of bacterial population sizes were similar on seedlings, and no defense responses were induced evidencing bacterial colonization of seedlings without any associated defense response induction. Bacteria associated with the spermosphere multiply in this rich environment, suggesting that the colonization of seedlings relies mostly on commensalism. The transmission of plant-pathogenic bacteria to and by nonhost seeds suggests a probable role of seeds of nonhost plants as an inoculum source.
Figures
References
-
- Ah-You, N., L. Gagnevin, P. A. D. Grimont, S. Brisse, X. Nesme, F. Chiroleu, L. B. T. Ngoc, E. Jouen, P. Lefeuvre, C. Vernière, and O. Pruvost. 2009. Polyphasic characterization of xanthomonads pathogenic to members of the Anacardiacae and their relatedness to the species of Xanthomonas. Int. J. Syst. Evol. Microbiol. 59:306-318. - PubMed
-
- Arlat, M., C. L. Gough, C. E. Barber, C. Boucher, and M. J. Daniels. 1991. Xanthomonas campestris contains a cluster of hrp genes related to the larger hrp cluster of Pseudomonas solanacearum. Mol. Plant Microbe Interact. 4:593-601. - PubMed
-
- Boureau, T., H. ElMaarouf-Bouteau, A. Garnier, M.-N. Brisset, C. Perino, I. Pucheu, and M.-A. Barny. 2006. DspA/E, a type III effector essential for Erwinia amylovora pathogenicity and growth in planta, induces cell death in host apple and non-host tobacco plants. Mol. Plant Microbe Interact. 19:16-24. - PubMed
-
- Bradford, M. M. 1976. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254. - PubMed
-
- Brunings, A. M., and D. W. Gabriel. 2003. Xanthomonas citri: breaking the surface. Mol. Plant Pathol. 4:141-157. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
