

Fold-change detection and scalar symmetry of sensory input fields
- PMID: 20729472
- PMCID: PMC2936624
- DOI: 10.1073/pnas.1002352107
Fold-change detection and scalar symmetry of sensory input fields
Abstract
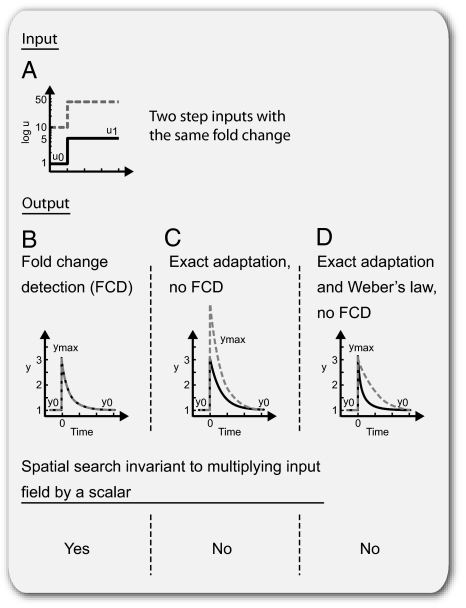
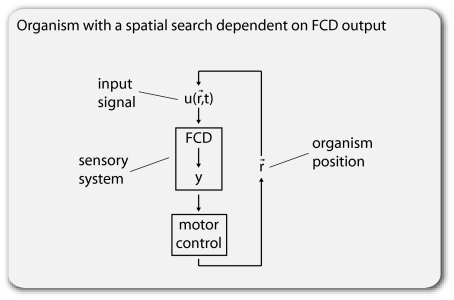
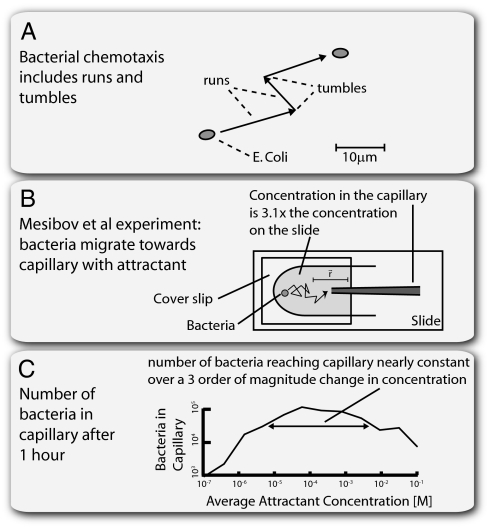
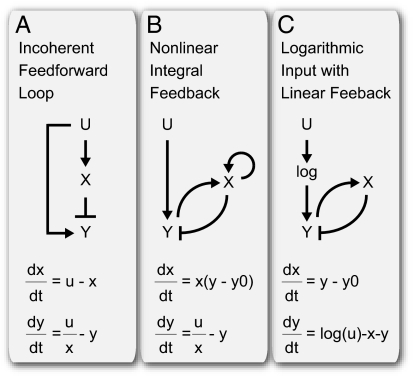
Recent studies suggest that certain cellular sensory systems display fold-change detection (FCD): a response whose entire shape, including amplitude and duration, depends only on fold changes in input and not on absolute levels. Thus, a step change in input from, for example, level 1 to 2 gives precisely the same dynamical output as a step from level 2 to 4, because the steps have the same fold change. We ask what the benefit of FCD is and show that FCD is necessary and sufficient for sensory search to be independent of multiplying the input field by a scalar. Thus, the FCD search pattern depends only on the spatial profile of the input and not on its amplitude. Such scalar symmetry occurs in a wide range of sensory inputs, such as source strength multiplying diffusing/convecting chemical fields sensed in chemotaxis, ambient light multiplying the contrast field in vision, and protein concentrations multiplying the output in cellular signaling systems. Furthermore, we show that FCD entails two features found across sensory systems, exact adaptation and Weber's law, but that these two features are not sufficient for FCD. Finally, we present a wide class of mechanisms that have FCD, including certain nonlinear feedback and feed-forward loops. We find that bacterial chemotaxis displays feedback within the present class and hence, is expected to show FCD. This can explain experiments in which chemotaxis searches are insensitive to attractant source levels. This study, thus, suggests a connection between properties of biological sensory systems and scalar symmetry stemming from physical properties of their input fields.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Response rescaling in bacterial chemotaxis.Proc Natl Acad Sci U S A. 2011 Aug 16;108(33):13870-5. doi: 10.1073/pnas.1108608108. Epub 2011 Aug 1. Proc Natl Acad Sci U S A. 2011. PMID: 21808031 Free PMC article.
-
Logarithmic and power law input-output relations in sensory systems with fold-change detection.PLoS Comput Biol. 2014 Aug 14;10(8):e1003781. doi: 10.1371/journal.pcbi.1003781. eCollection 2014 Aug. PLoS Comput Biol. 2014. PMID: 25121598 Free PMC article.
-
Cellular sensory mechanisms for detecting specific fold-changes in extracellular cues.Biophys J. 2014 Jan 7;106(1):279-88. doi: 10.1016/j.bpj.2013.10.039. Biophys J. 2014. PMID: 24411260 Free PMC article.
-
Input-output relations in biological systems: measurement, information and the Hill equation.Biol Direct. 2013 Dec 5;8:31. doi: 10.1186/1745-6150-8-31. Biol Direct. 2013. PMID: 24308849 Free PMC article. Review.
-
Human brain plasticity: evidence from sensory deprivation and altered language experience.Prog Brain Res. 2002;138:177-88. doi: 10.1016/S0079-6123(02)38078-6. Prog Brain Res. 2002. PMID: 12432770 Review.
Cited by
-
Background compensation revisited: Conserved phase response curves in frequency controlled homeostats with coherent feedback.PLoS One. 2024 Sep 4;19(9):e0305804. doi: 10.1371/journal.pone.0305804. eCollection 2024. PLoS One. 2024. PMID: 39231133 Free PMC article.
-
Chemotactic smoothing of collective migration.Elife. 2022 Mar 8;11:e71226. doi: 10.7554/eLife.71226. Elife. 2022. PMID: 35257660 Free PMC article.
-
Response rescaling in bacterial chemotaxis.Proc Natl Acad Sci U S A. 2011 Aug 16;108(33):13870-5. doi: 10.1073/pnas.1108608108. Epub 2011 Aug 1. Proc Natl Acad Sci U S A. 2011. PMID: 21808031 Free PMC article.
-
Optimality principles in the regulation of metabolic networks.Metabolites. 2012 Aug 29;2(3):529-52. doi: 10.3390/metabo2030529. Metabolites. 2012. PMID: 24957646 Free PMC article.
-
Responding to chemical gradients: bacterial chemotaxis.Curr Opin Cell Biol. 2012 Apr;24(2):262-8. doi: 10.1016/j.ceb.2011.11.008. Epub 2011 Dec 9. Curr Opin Cell Biol. 2012. PMID: 22169400 Free PMC article. Review.
References
-
- Kandel ER, Schwartz JH, Jessell TM. Principles of Neural Science. 4th Ed. New York: McGraw-Hill; 2000.
-
- Smirnakis SM, Berry MJ, Warland DK, Bialek W, Meister M. Adaptation of retinal processing to image contrast and spatial scale. Nature. 1997;386:69–73. - PubMed
-
- Keener J, Sneyd J. Mathematical Physiology. 2nd Ed. Berlin: Springer; 2009.
-
- Weber EH. Tatsinn and Gemeingefuhl. Leipzig, Germany: Verlag von Wilhelm Englemann; 1905.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
