

TRIM16 acts as a tumour suppressor by inhibitory effects on cytoplasmic vimentin and nuclear E2F1 in neuroblastoma cells
- PMID: 20729920
- PMCID: PMC3007621
- DOI: 10.1038/onc.2010.340
TRIM16 acts as a tumour suppressor by inhibitory effects on cytoplasmic vimentin and nuclear E2F1 in neuroblastoma cells
Abstract
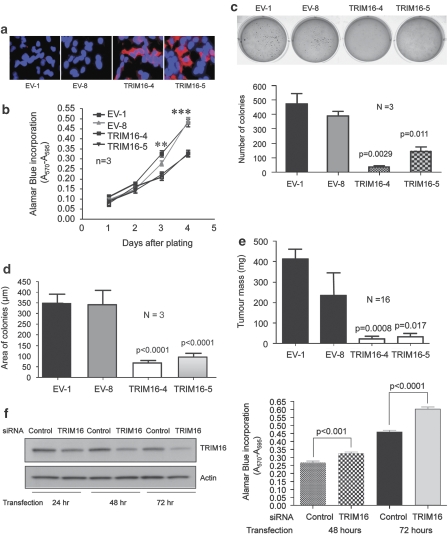
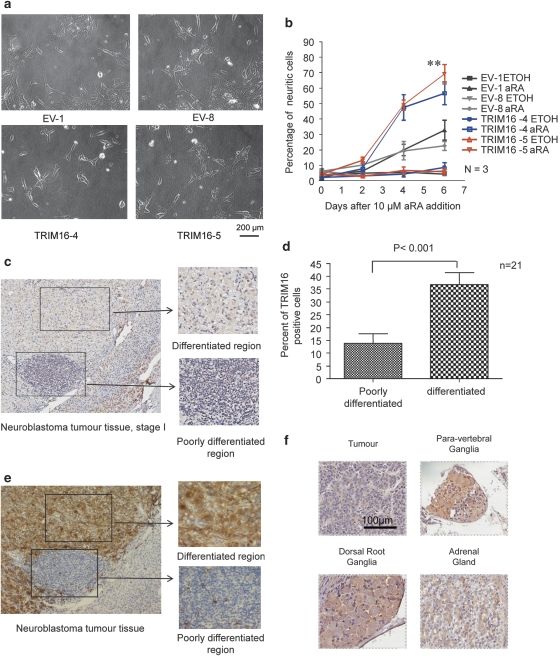
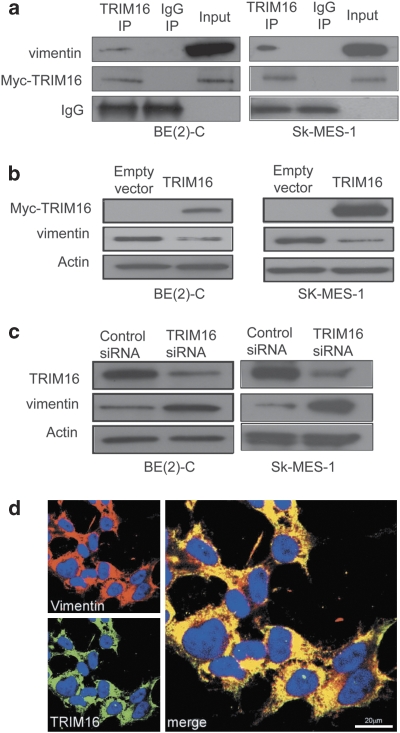
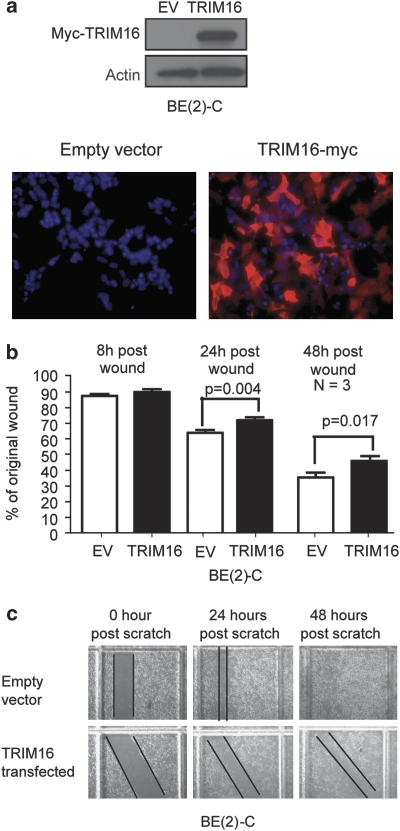
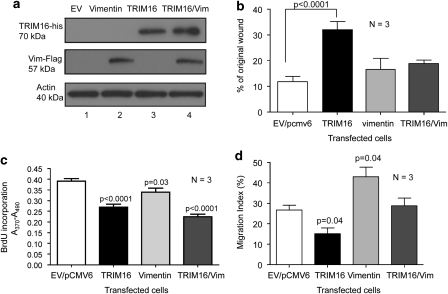
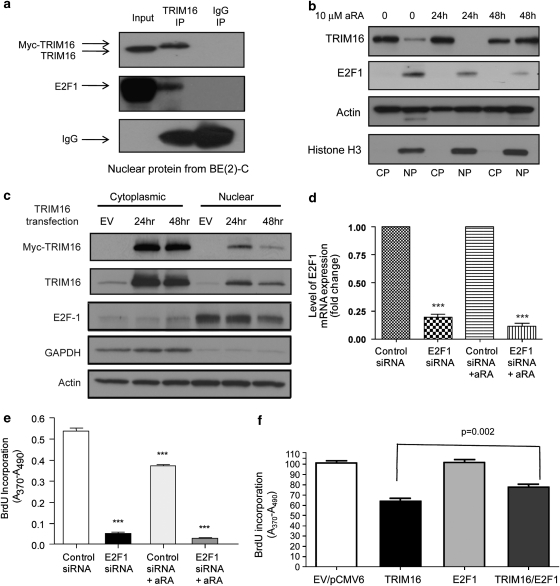
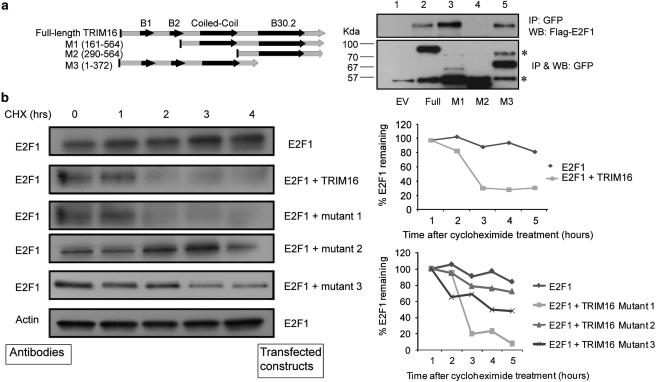
The family of tripartite-motif (TRIM) proteins are involved in diverse cellular processes, but are often characterized by critical protein-protein interactions necessary for their function. TRIM16 is induced in different cancer types, when the cancer cell is forced to proceed down a differentiation pathway. We have identified TRIM16 as a DNA-binding protein with histone acetylase activity, which is required for the retinoic acid receptor β(2) transcriptional response in retinoid-treated cancer cells. In this study, we show that overexpressed TRIM16 reduced neuroblastoma cell growth, enhanced retinoid-induced differentiation and reduced tumourigenicity in vivo. TRIM16 was only expressed in the differentiated ganglion cell component of primary human neuroblastoma tumour tissues. TRIM16 bound directly to cytoplasmic vimentin and nuclear E2F1 in neuroblastoma cells. TRIM16 reduced cell motility and this required downregulation of vimentin. Retinoid treatment and enforced overexpression caused TRIM16 to translocate to the nucleus, and bind to and downregulate nuclear E2F1, required for cell replication. This study, for the first time, demonstrates that TRIM16 acts as a tumour suppressor, affecting neuritic differentiation, cell migration and replication through interactions with cytoplasmic vimentin and nuclear E2F1 in neuroblastoma cells.
Figures
Similar articles
-
The retinoid signalling molecule, TRIM16, is repressed during squamous cell carcinoma skin carcinogenesis in vivo and reduces skin cancer cell migration in vitro.J Pathol. 2012 Feb;226(3):451-62. doi: 10.1002/path.2986. Epub 2011 Oct 18. J Pathol. 2012. PMID: 22009481 Free PMC article.
-
High TDP43 expression is required for TRIM16-induced inhibition of cancer cell growth and correlated with good prognosis of neuroblastoma and breast cancer patients.Cancer Lett. 2016 May 1;374(2):315-23. doi: 10.1016/j.canlet.2016.02.021. Epub 2016 Feb 20. Cancer Lett. 2016. PMID: 26902425
-
TRIM16 inhibits neuroblastoma cell proliferation through cell cycle regulation and dynamic nuclear localization.Cell Cycle. 2013 Mar 15;12(6):889-98. doi: 10.4161/cc.23825. Epub 2013 Feb 19. Cell Cycle. 2013. PMID: 23422002 Free PMC article.
-
TRIM proteins in neuroblastoma.Biosci Rep. 2019 Dec 20;39(12):BSR20192050. doi: 10.1042/BSR20192050. Biosci Rep. 2019. PMID: 31820796 Free PMC article. Review.
-
Negative autoregulation of p73 and p53 by DeltaNp73 in regulating differentiation and survival of human neuroblastoma cells.Cancer Lett. 2003 Jul 18;197(1-2):105-9. doi: 10.1016/s0304-3835(03)00090-9. Cancer Lett. 2003. PMID: 12880968 Review.
Cited by
-
Changes in the Expression of Long Non-Coding RNA SDMGC and Its Target Gene, TRIM16, in Patients with Gastric Cancer.J Gastrointest Cancer. 2023 Mar;54(1):44-50. doi: 10.1007/s12029-021-00791-y. Epub 2022 Jan 3. J Gastrointest Cancer. 2023. PMID: 34978663
-
TRIM59 Promotes the Proliferation and Migration of Non-Small Cell Lung Cancer Cells by Upregulating Cell Cycle Related Proteins.PLoS One. 2015 Nov 24;10(11):e0142596. doi: 10.1371/journal.pone.0142596. eCollection 2015. PLoS One. 2015. PMID: 26599082 Free PMC article.
-
Unveiling the intersection: ferroptosis in influenza virus infection.Virol J. 2024 Aug 12;21(1):185. doi: 10.1186/s12985-024-02462-3. Virol J. 2024. PMID: 39135112 Free PMC article. Review.
-
Thymosin-β4 is a determinant of drug sensitivity for Fenretinide and Vorinostat combination therapy in neuroblastoma.Mol Oncol. 2015 Aug;9(7):1484-500. doi: 10.1016/j.molonc.2015.04.005. Epub 2015 Apr 29. Mol Oncol. 2015. PMID: 25963741 Free PMC article.
-
Withaferin A-A Promising Phytochemical Compound with Multiple Results in Dermatological Diseases.Molecules. 2021 Apr 21;26(9):2407. doi: 10.3390/molecules26092407. Molecules. 2021. PMID: 33919088 Free PMC article. Review.
References
-
- Beer HD, Munding C, Dubois N, Mamie C, Hohl D, Werner S. The estrogen-responsive B box protein: a novel regulator of keratinocyte differentiation. J Biol Chem. 2002;277:20740–20749. - PubMed
-
- Borden KL, Martin SR, O'Reilly NJ, Lally JM, Reddy BA, Etkin LD, et al. Characterisation of a novel cysteine/histidine-rich metal binding domain from Xenopus nuclear factor XNF7. FEBS Letters. 1993;335:255–260. - PubMed
-
- Cao T, Duprez E, Borden KL, Freemont PS, Etkin LD. Ret finger protein is a normal component of PML nuclear bodies and interacts directly with PML. J Cell Sci. 1998;111:1319–1329. - PubMed
-
- Chae JJ, Komarow HD, Cheng J, Wood G, Raben N, Liu PP, et al. Targeted disruption of pyrin, the FMF protein, causes heightened sensitivity to endotoxin and a defect in macrophage apoptosis. Mol Cell. 2003;11:591–604. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
