

Review
doi: 10.1038/nrm2957.
Cell adhesion: integrating cytoskeletal dynamics and cellular tension
Affiliations
- PMID: 20729930
- PMCID: PMC2992881
- DOI: 10.1038/nrm2957
Item in Clipboard
Review
Cell adhesion: integrating cytoskeletal dynamics and cellular tension
Nat Rev Mol Cell Biol.
2010 Sep.
Abstract
Cell migration affects all morphogenetic processes and contributes to numerous diseases, including cancer and cardiovascular disease. For most cells in most environments, movement begins with protrusion of the cell membrane followed by the formation of new adhesions at the cell front that link the actin cytoskeleton to the substratum, generation of traction forces that move the cell forwards and disassembly of adhesions at the cell rear. Adhesion formation and disassembly drive the migration cycle by activating Rho GTPases, which in turn regulate actin polymerization and myosin II activity, and therefore adhesion dynamics.
Figures
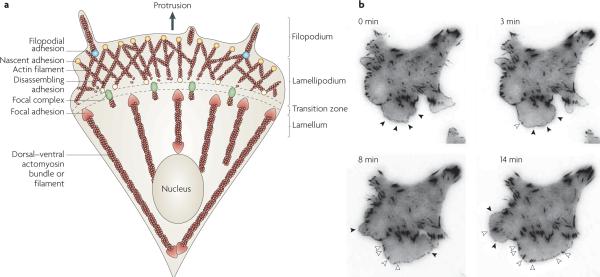
a | Adhesion is closely coupled with the protrusions of the leading edge of the cell (filopodia and lamellipodia). Adhesions (nascent adhesions) initially form in the lamellipodium (although adhesions may also be associated with filopodia) and the rate of nascent adhesion assembly correlates with the rate of protrusion. Nascent adhesions either disassemble or elongate at the convergence of the lamellipodium and lamellum (the transition zone). Adhesion maturation to focal complexes and focal adhesions is accompanied by the bundling and cross-bridging of actin filaments, and actomyosin-induced contractility stabilizes adhesion formation and increases adhesion size. b | TIRF micrographs of a Chinese hamster ovary (CHO) cell expressing paxillin–mEGFP (monomeric enhanced green fluorescent protein) on glass coated with fibronectin (5 μg ml−1). Images were acquired every 5 seconds, and representative images from 0, 3, 8 and 14 minutes are shown (see REF. 49). Closed arrow heads denote nascent adhesions assembling and turning over in protrusions. Open arrow heads indicate maturing adhesions that begin to elongate centripetally (that is, towards the cell centre) when protrusion pauses or halts. For a movie of this experiment see supplementary information S1 (movie).
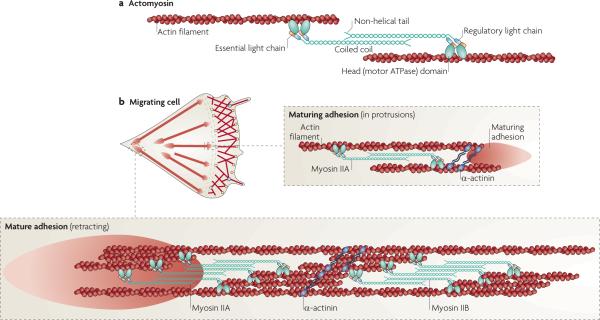
a | Adhesions elongate along actin filaments that contain myosin IIA, which cross links the actin filaments and exerts tension on them. This leads to tension on the conformational sensitivity, and clustering of, adhesion molecules that are directly or indirectly associated with actin. Myosin II activity is regulated by phosphorylation on the regulatory light chain at Thr18 and Ser19, although other regulatory sites in the heavy chain are also implicated in its activities. For a more complete discussion of myosin II structure and function see REF. 8. b | In a migrating cell, myosin IIA acts at a distance to regulate adhesion maturation and turnover as it is juxtaposed to, but not directly associated with, the maturing adhesion at the cell front. α-actinin cross links actin filaments. Adhesions at the rear are associated with large actin filament bundles that contain both myosin IIA and myosin IIB. Their activity mediates rear retraction and adhesion disassembly.
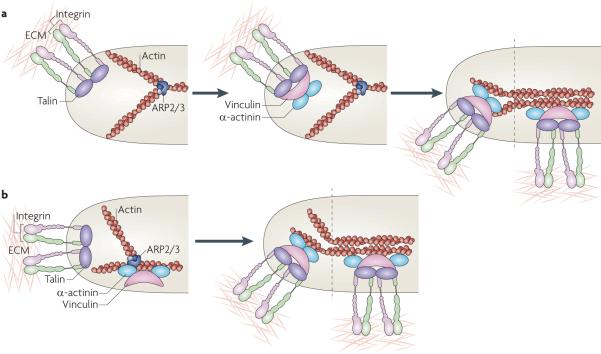
a | In one model, adhesion nucleation is initiated by the binding of integrins to extracellular matrix (ECM) proteins, their ligand-mediated clustering and the coordinate assembly of new adhesion complexes on the clustered integrin cytoplasmic domains, which are depicted here as a complex with talin, vinculin, α-actinin and dendritic actin (middle panel). Maturation of the adhesions is mediated by increased tension on them and the bundling and cross-bridging of the actin filaments (right panel). b | A second model posits that adhesion formation is coupled to actin polymerization and that vinculin (and perhaps focal adhesion kinase (FAK)) bind directly to actin-related protein 2/3 (ARP2/3) complexes and colocalize before adhesion formation (left panel). These complexes then bind integrins (depicted here in association with talin), stabilizing the nascent adhesion (right panel). As in part a, maturation of the adhesions is mediated by increased tension on the adhesions and the bundling and cross-bridging of the actin filaments.
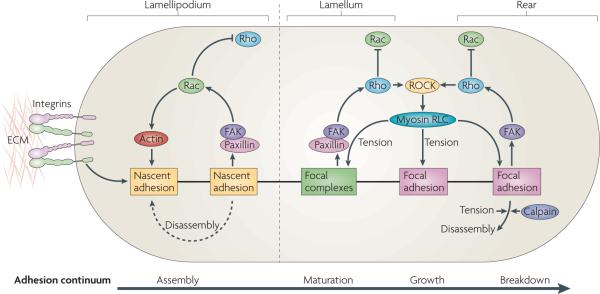
Nascent adhesion formation and disassembly are coupled with the forward movement of the lamellipodium. Maturation of adhesions is dependent on actomyosin in the lamellum, where adhesions become larger. Adhesion formation and disassembly in the lamellipodium is driven by the activation of Rac (and perhaps the localized suppression of Rho activity), which involves activation of the Tyr-phosphorylated scaffolds, paxillin and focal adhesion kinase (FAK). In the lamellum, adhesion maturation is accompanied by localized activation of Rho, perhaps through FAK-dependent recruitment of Rho guanine nucleotide exchange factors (GEFs) and Rho GTPase-activating proteins (GAPs). Rho activation sustains the activation of myosin II through the action of Rho-associated protein kinase (ROCK), which controls the kinases and phosphatases that regulate its regulatory light chain (RLC) phosphorylation. Myosin II-generated tension sustains adhesion maturation by cross linking- and tension-induced conformational changes in various adhesion proteins (see main text). Disassembly of adhesions at the cell rear is Rho GTPase- and myosin II-dependent, and may also involve the action of proteases, such as calpains, on adhesion-linked proteins. ECM, extracellular matrix.
References
-
- Campbell ID, Ginsberg MH. The talin–tail interaction places integrin activation on FERM ground. Trends Biochem. Sci. 2004;29:429–435. - PubMed
-
- Premont RT. The GIT/PIX complex: an oligomeric assembly of GIT family ARF GTPase-activating proteins and PIX family Rac1/Cdc42 guanine nucleotide exchange factors. Cell. Signal. 2004;16:1001–1011. - PubMed
-
- Geiger B, Spatz JP, Bershadsky AD. Environmental sensing through focal adhesions. Nature Rev. Mol. Cell Biol. 2009;10:21–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
