

Open source tools for the information theoretic analysis of neural data
- PMID: 20730105
- PMCID: PMC2891486
- DOI: 10.3389/neuro.01.011.2010
Open source tools for the information theoretic analysis of neural data
Abstract
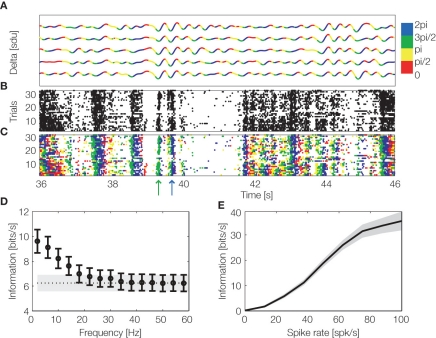
The recent and rapid development of open source software tools for the analysis of neurophysiological datasets consisting of simultaneous multiple recordings of spikes, field potentials and other neural signals holds the promise for a significant advance in the standardization, transparency, quality, reproducibility and variety of techniques used to analyze neurophysiological data and for the integration of information obtained at different spatial and temporal scales. In this review we focus on recent advances in open source toolboxes for the information theoretic analysis of neural responses. We also present examples of their use to investigate the role of spike timing precision, correlations across neurons, and field potential fluctuations in the encoding of sensory information. These information toolboxes, available both in MATLAB and Python programming environments, hold the potential to enlarge the domain of application of information theory to neuroscience and to lead to new discoveries about how neurons encode and transmit information.
Keywords: bias; entropy; information theory; mutual information; open source.
Figures
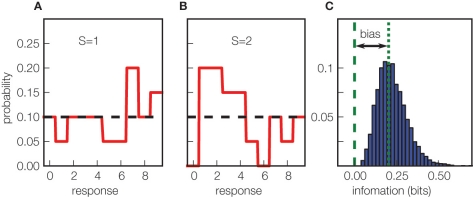
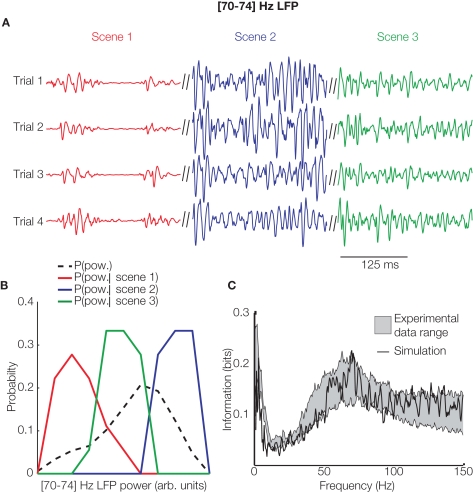
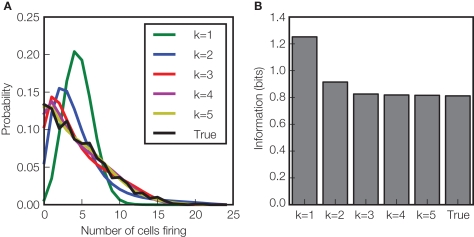
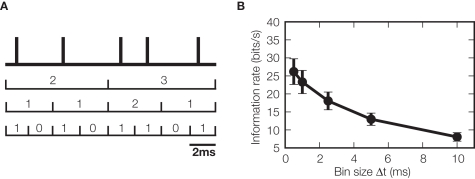
References
-
- Adrian E. D. (1928). The Basis of Sensation. New York, Norton
Grants and funding
LinkOut - more resources
Full Text Sources