

Uniting sex and eukaryote origins in an emerging oxygenic world
- PMID: 20731852
- PMCID: PMC2933680
- DOI: 10.1186/1745-6150-5-53
Uniting sex and eukaryote origins in an emerging oxygenic world
Abstract
Background: Theories about eukaryote origins (eukaryogenesis) need to provide unified explanations for the emergence of diverse complex features that define this lineage. Models that propose a prokaryote-to-eukaryote transition are gridlocked between the opposing "phagocytosis first" and "mitochondria as seed" paradigms, neither of which fully explain the origins of eukaryote cell complexity. Sex (outcrossing with meiosis) is an example of an elaborate trait not yet satisfactorily addressed in theories about eukaryogenesis. The ancestral nature of meiosis and its dependence on eukaryote cell biology suggest that the emergence of sex and eukaryogenesis were simultaneous and synergic and may be explained by a common selective pressure.
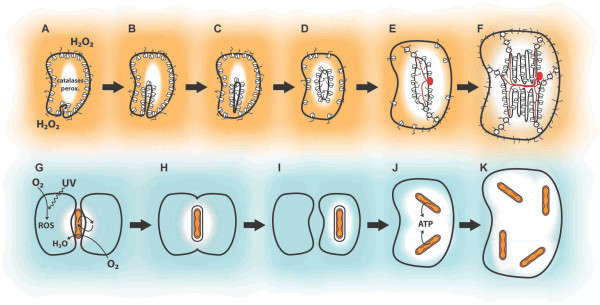
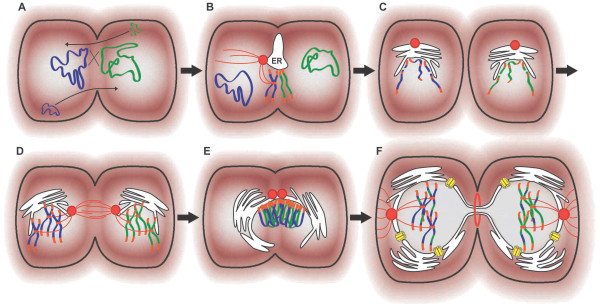
Presentation of the hypothesis: We propose that a local rise in oxygen levels, due to cyanobacterial photosynthesis in ancient Archean microenvironments, was highly toxic to the surrounding biota. This selective pressure drove the transformation of an archaeal (archaebacterial) lineage into the first eukaryotes. Key is that oxygen might have acted in synergy with environmental stresses such as ultraviolet (UV) radiation and/or desiccation that resulted in the accumulation of reactive oxygen species (ROS). The emergence of eukaryote features such as the endomembrane system and acquisition of the mitochondrion are posited as strategies to cope with a metabolic crisis in the cell plasma membrane and the accumulation of ROS, respectively. Selective pressure for efficient repair of ROS/UV-damaged DNA drove the evolution of sex, which required cell-cell fusions, cytoskeleton-mediated chromosome movement, and emergence of the nuclear envelope. Our model implies that evolution of sex and eukaryogenesis were inseparable processes.
Testing the hypothesis: Several types of data can be used to test our hypothesis. These include paleontological predictions, simulation of ancient oxygenic microenvironments, and cell biological experiments with Archaea exposed to ROS and UV stresses. Studies of archaeal conjugation, prokaryotic DNA recombination, and the universality of nuclear-mediated meiotic activities might corroborate the hypothesis that sex and the nucleus evolved to support DNA repair.
Implications of the hypothesis: Oxygen tolerance emerges as an important principle to investigate eukaryogenesis. The evolution of eukaryotic complexity might be best understood as a synergic process between key evolutionary innovations, of which meiosis (sex) played a central role.
Reviewers: This manuscript was reviewed by Eugene V. Koonin, Anthony M. Poole, and Gáspár Jékely.
Figures
Similar articles
-
Mitochondria, the Cell Cycle, and the Origin of Sex via a Syncytial Eukaryote Common Ancestor.Genome Biol Evol. 2016 Jul 2;8(6):1950-70. doi: 10.1093/gbe/evw136. Genome Biol Evol. 2016. PMID: 27345956 Free PMC article.
-
The viral eukaryogenesis hypothesis: a key role for viruses in the emergence of eukaryotes from a prokaryotic world environment.Ann N Y Acad Sci. 2009 Oct;1178:91-105. doi: 10.1111/j.1749-6632.2009.04994.x. Ann N Y Acad Sci. 2009. PMID: 19845630
-
The emerging view on the origin and early evolution of eukaryotic cells.Nature. 2024 Sep;633(8029):295-305. doi: 10.1038/s41586-024-07677-6. Epub 2024 Sep 11. Nature. 2024. PMID: 39261613 Review.
-
Coevolution of Eukaryote-like Vps4 and ESCRT-III Subunits in the Asgard Archaea.mBio. 2020 May 19;11(3):e00417-20. doi: 10.1128/mBio.00417-20. mBio. 2020. PMID: 32430468 Free PMC article.
-
Endosymbiotic selective pressure at the origin of eukaryotic cell biology.Elife. 2022 Nov 10;11:e81033. doi: 10.7554/eLife.81033. Elife. 2022. PMID: 36355038 Free PMC article. Review.
Cited by
-
Sex determination strategies in 2012: towards a common regulatory model?Reprod Biol Endocrinol. 2012 Feb 22;10:13. doi: 10.1186/1477-7827-10-13. Reprod Biol Endocrinol. 2012. PMID: 22357269 Free PMC article. Review.
-
Integrated conjugative plasmid drives high frequency chromosomal gene transfer in Sulfolobus islandicus.Front Microbiol. 2023 Jan 23;14:1114574. doi: 10.3389/fmicb.2023.1114574. eCollection 2023. Front Microbiol. 2023. PMID: 36756353 Free PMC article.
-
Towards a processual microbial ontology.Biol Philos. 2013 Mar;28(2):379-404. doi: 10.1007/s10539-012-9350-2. Epub 2012 Nov 6. Biol Philos. 2013. PMID: 23487350 Free PMC article.
-
Primal eukaryogenesis: on the communal nature of precellular States, ancestral to modern life.Life (Basel). 2012 Jan 23;2(1):170-212. doi: 10.3390/life2010170. Life (Basel). 2012. PMID: 25382122 Free PMC article.
-
Origins of eukaryotic sexual reproduction.Cold Spring Harb Perspect Biol. 2014 Mar 1;6(3):a016154. doi: 10.1101/cshperspect.a016154. Cold Spring Harb Perspect Biol. 2014. PMID: 24591519 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
