

Allelic diversity and naturally acquired allele-specific antibody responses to Plasmodium falciparum apical membrane antigen 1 in Kenya
- PMID: 20732997
- PMCID: PMC2976345
- DOI: 10.1128/IAI.00576-10
Allelic diversity and naturally acquired allele-specific antibody responses to Plasmodium falciparum apical membrane antigen 1 in Kenya
Abstract
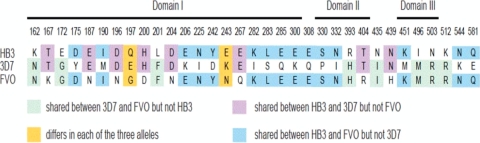
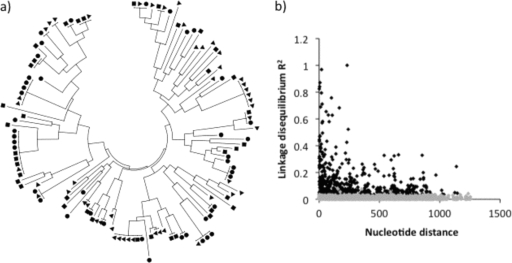
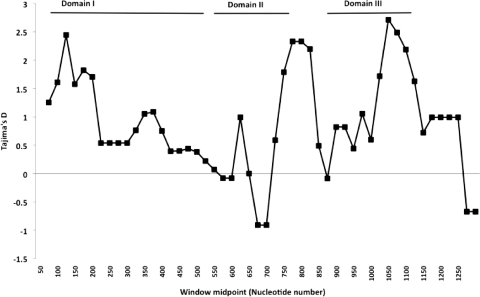
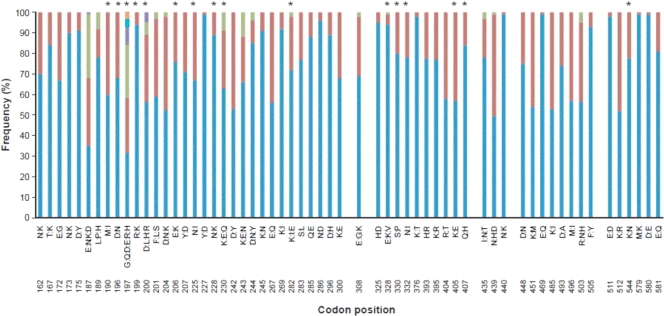
Although Plasmodium falciparum apical membrane antigen 1 (AMA1) is a leading malaria vaccine candidate, extensive allelic diversity may compromise its vaccine potential. We have previously shown that naturally acquired antibodies to AMA1 were associated with protection from clinical malaria in this Kenyan population. To assess the impact of allelic diversity on naturally acquired immunity, we first sequenced the ectodomain-encoding region of P. falciparum ama1 from subjects with asymptomatic, mild, and severe malaria and measured allele frequency distributions. We then measured antibodies to three allelic AMA1 proteins (AMA1_3D7, AMA1_FVO, and AMA1_HB3) and used competition enzyme-linked immunosorbent assays (ELISAs) to analyze allele-specific antibodies. Seventy-eight unique haplotypes were identified from 129 alleles sampled. No clustering of allelic haplotypes with disease severity or year of sampling was observed. Differences in nucleotide frequencies in clinical (severe plus mild malaria) versus asymptomatic infections were observed at 16 polymorphic positions. Allele frequency distributions were indicative of balancing selection, with the strongest signature being identified in domain III (Tajima's D = 2.51; P < 0.05). Antibody reactivities to each of the three allelic AMA1 proteins were highly correlated (P < 0.001 for all pairwise comparisons). Although antibodies to conserved epitopes were abundant, 48% of selected children with anti-AMA1 IgG (n = 106) had detectable reactivity to allele-specific epitopes as determined by a competition ELISA. Antibodies to both conserved and allele-specific epitopes in AMA1 may contribute to clinical protection.
Figures
References
-
- Anders, R. F., P. E. Crewther, S. Edwards, M. Margetts, M. L. Matthew, B. Pollock, and D. Pye. 1998. Immunisation with recombinant AMA-1 protects mice against infection with Plasmodium chabaudi. Vaccine 16:240-247. - PubMed
-
- Babiker, H. A., J. Lines, W. G. Hill, and D. Walliker. 1997. Population structure of Plasmodium falciparum in villages with different malaria endemicity in east Africa. Am. J. Trop. Med. Hyg. 56:141-147. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
