

How cyclin A destruction escapes the spindle assembly checkpoint
- PMID: 20733051
- PMCID: PMC2928024
- DOI: 10.1083/jcb.201001083
How cyclin A destruction escapes the spindle assembly checkpoint
Abstract
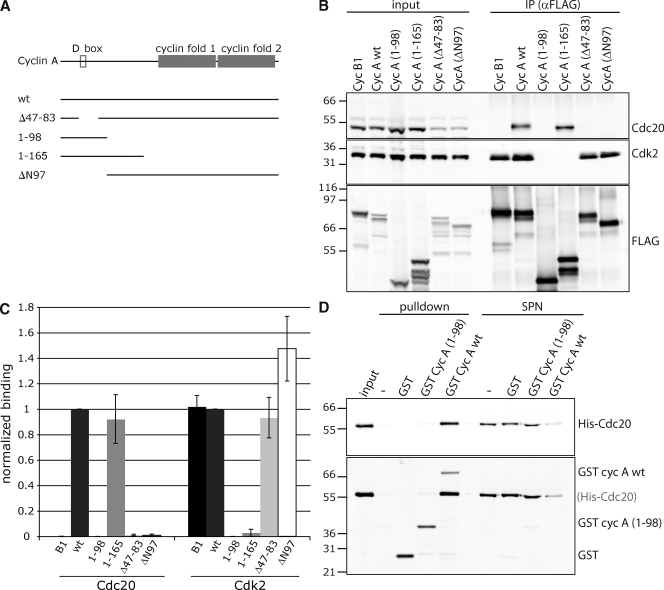
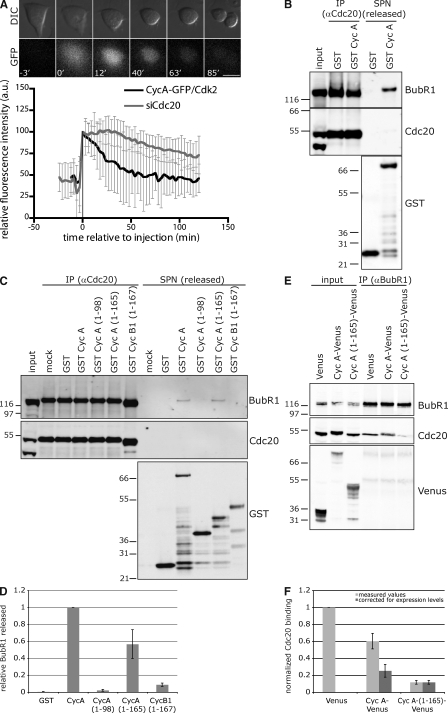
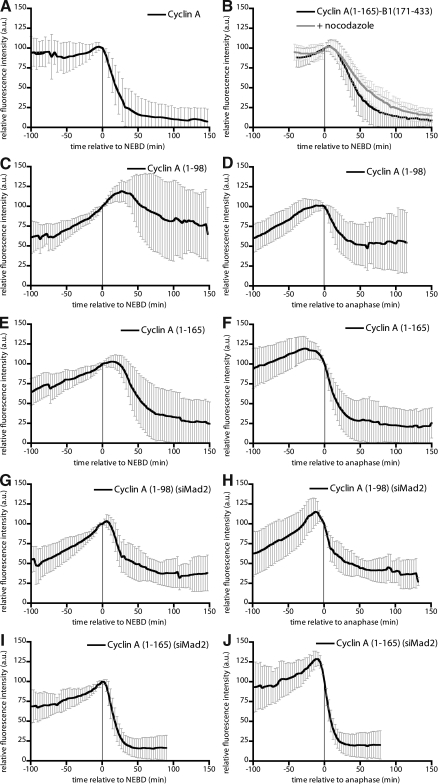
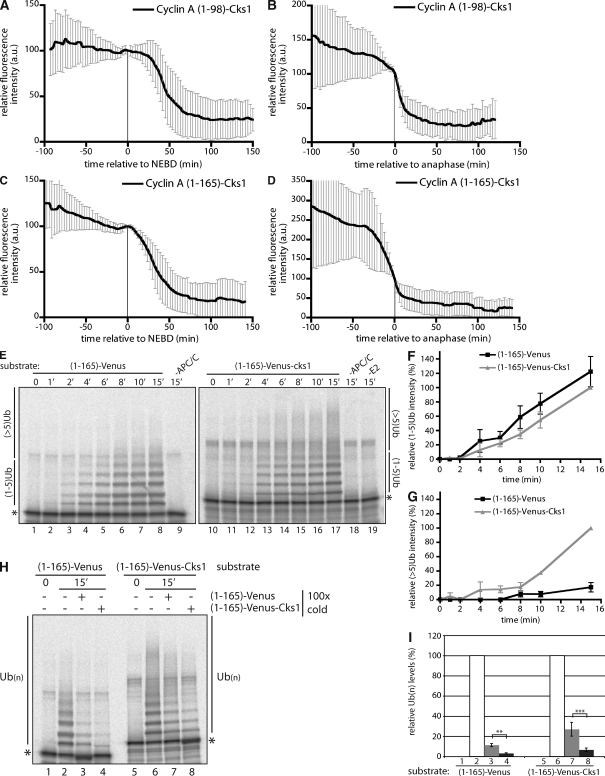
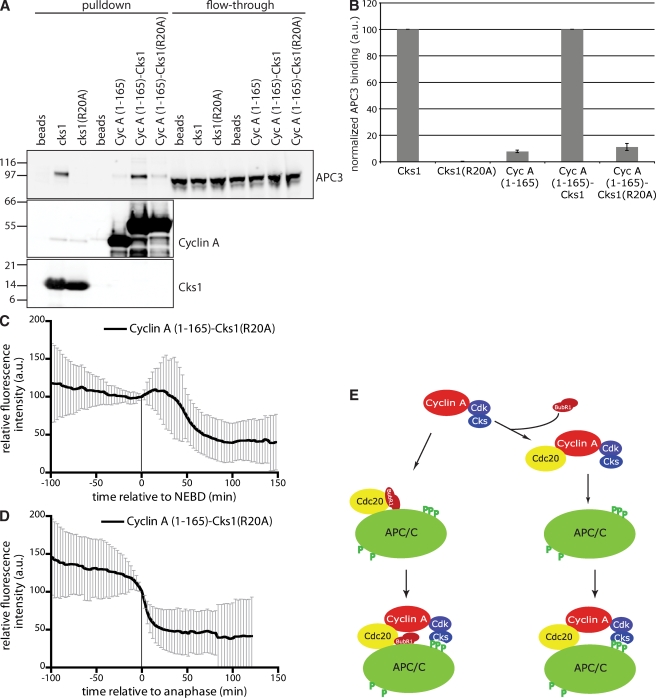
The anaphase-promoting complex/cyclosome (APC/C) is the ubiquitin ligase essential to mitosis, which ensures that specific proteins are degraded at specific times to control the order of mitotic events. The APC/C coactivator, Cdc20, is targeted by the spindle assembly checkpoint (SAC) to restrict APC/C activity until metaphase, yet early substrates, such as cyclin A, are degraded in the presence of the active checkpoint. Cdc20 and the cyclin-dependent kinase cofactor, Cks, are required for cyclin A destruction, but how they enable checkpoint-resistant destruction has not been elucidated. In this study, we answer this problem: we show that the N terminus of cyclin A binds directly to Cdc20 and with sufficient affinity that it can outcompete the SAC proteins. Subsequently, the Cks protein is necessary and sufficient to promote cyclin A degradation in the presence of an active checkpoint by binding cyclin A-Cdc20 to the APC/C.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
