

Neuropeptidomic components generated by proteomic functions in secretory vesicles for cell-cell communication
- PMID: 20734175
- PMCID: PMC2976990
- DOI: 10.1208/s12248-010-9223-z
Neuropeptidomic components generated by proteomic functions in secretory vesicles for cell-cell communication
Abstract
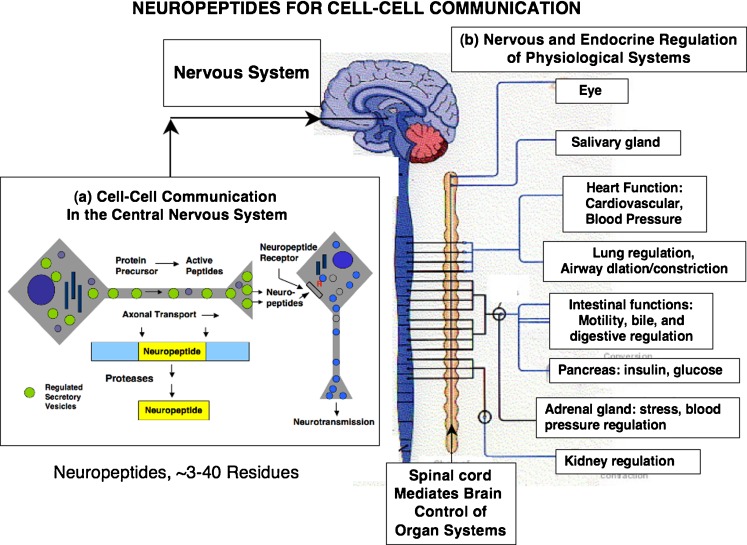
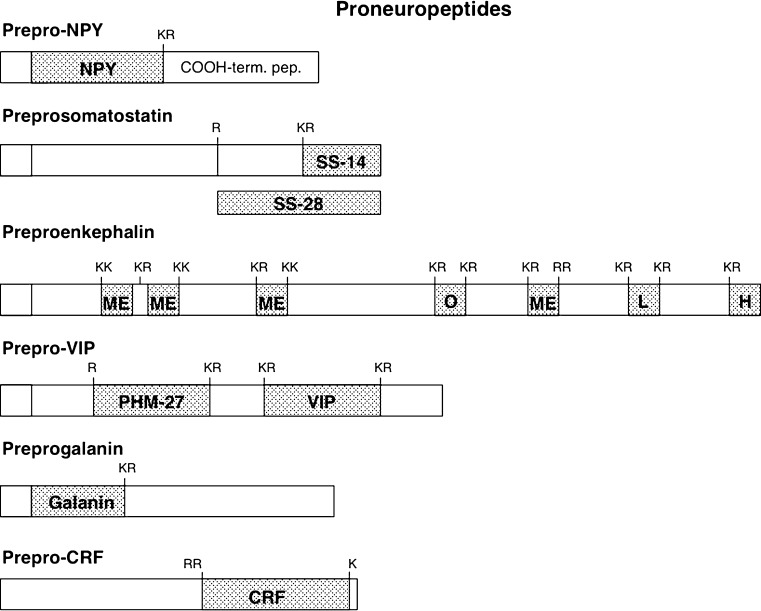
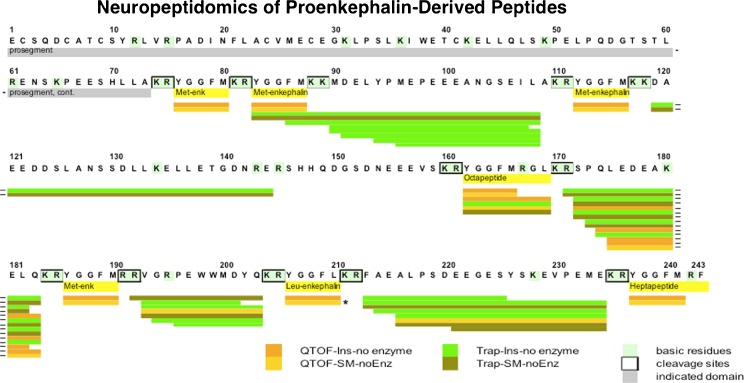
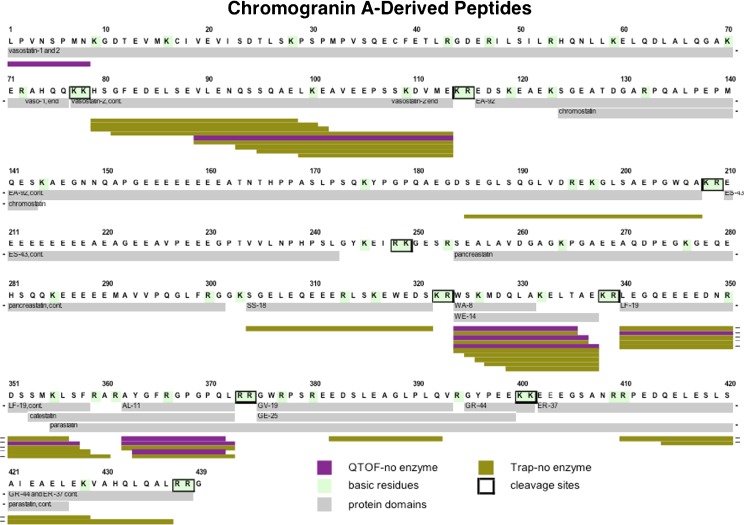
Diverse neuropeptides participate in cell-cell communication to coordinate neuronal and endocrine regulation of physiological processes in health and disease. Neuropeptides are short peptides ranging in length from ~3 to 40 amino acid residues that are involved in biological functions of pain, stress, obesity, hypertension, mental disorders, cancer, and numerous health conditions. The unique neuropeptide sequences define their specific biological actions. Significantly, this review article discusses how the neuropeptide field is at the crest of expanding knowledge gained from mass-spectrometry-based neuropeptidomic studies, combined with proteomic analyses for understanding the biosynthesis of neuropeptidomes. The ongoing expansion in neuropeptide diversity lies in the unbiased and global mass-spectrometry-based approaches for identification and quantitation of peptides. Current mass spectrometry technology allows definition of neuropeptide amino acid sequence structures, profiling of multiple neuropeptides in normal and disease conditions, and quantitative peptide measures in biomarker applications to monitor therapeutic drug efficacies. Complementary proteomic studies of neuropeptide secretory vesicles provide valuable insight into the protein processes utilized for neuropeptide production, storage, and secretion. Furthermore, ongoing research in developing new computational tools will facilitate advancements in mass-spectrometry-based identification of small peptides. Knowledge of the entire repertoire of neuropeptides that regulate physiological systems will provide novel insight into regulatory mechanisms in health, disease, and therapeutics.
Figures
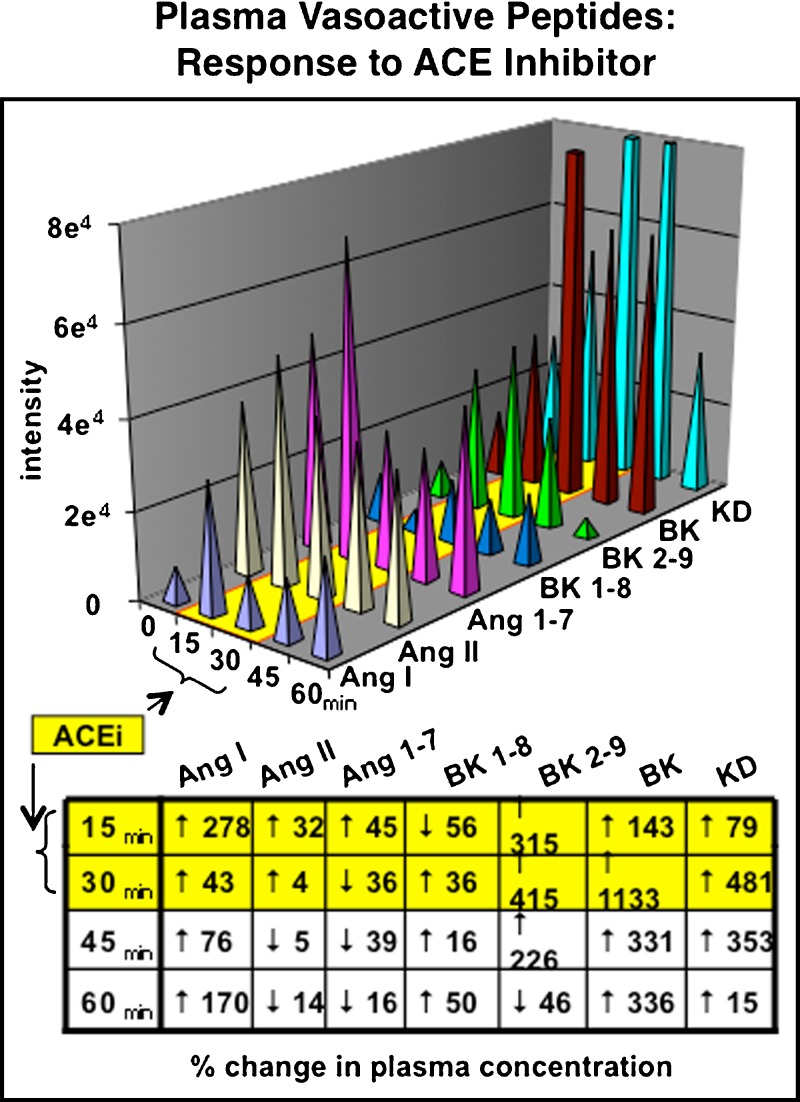
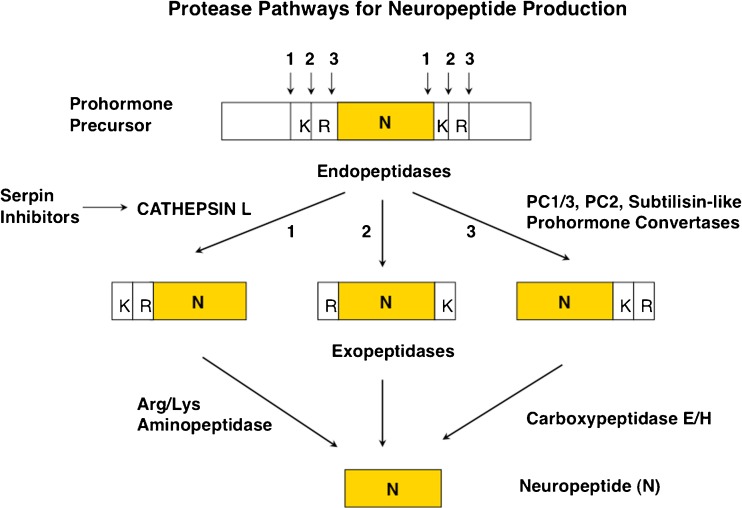
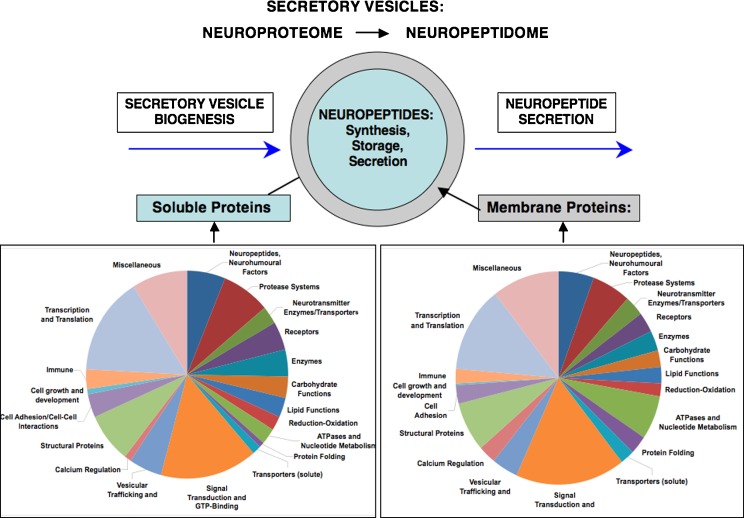
Similar articles
-
Neuropeptidomics Mass Spectrometry Reveals Signaling Networks Generated by Distinct Protease Pathways in Human Systems.J Am Soc Mass Spectrom. 2015 Dec;26(12):1970-80. doi: 10.1007/s13361-015-1251-6. Epub 2015 Oct 19. J Am Soc Mass Spectrom. 2015. PMID: 26483184 Free PMC article. Review.
-
Mass spectrometry-based neuropeptidomics of secretory vesicles from human adrenal medullary pheochromocytoma reveals novel peptide products of prohormone processing.J Proteome Res. 2010 Oct 1;9(10):5065-75. doi: 10.1021/pr100358b. J Proteome Res. 2010. PMID: 20704348 Free PMC article.
-
The novel role of cathepsin L for neuropeptide production illustrated by research strategies in chemical biology with protease gene knockout and expression.Methods Mol Biol. 2011;768:107-25. doi: 10.1007/978-1-61779-204-5_5. Methods Mol Biol. 2011. PMID: 21805239
-
Diversity of Neuropeptide Cell-Cell Signaling Molecules Generated by Proteolytic Processing Revealed by Neuropeptidomics Mass Spectrometry.J Am Soc Mass Spectrom. 2018 May;29(5):807-816. doi: 10.1007/s13361-018-1914-1. Epub 2018 Apr 17. J Am Soc Mass Spectrom. 2018. PMID: 29667161 Free PMC article. Review.
-
Unique biological function of cathepsin L in secretory vesicles for biosynthesis of neuropeptides.Neuropeptides. 2010 Dec;44(6):457-66. doi: 10.1016/j.npep.2010.08.003. Epub 2010 Nov 2. Neuropeptides. 2010. PMID: 21047684 Free PMC article. Review.
Cited by
-
NeuroPedia: neuropeptide database and spectral library.Bioinformatics. 2011 Oct 1;27(19):2772-3. doi: 10.1093/bioinformatics/btr445. Epub 2011 Aug 5. Bioinformatics. 2011. PMID: 21821666 Free PMC article.
-
The novel chromogranin A-derived serpinin and pyroglutaminated serpinin peptides are positive cardiac β-adrenergic-like inotropes.FASEB J. 2012 Jul;26(7):2888-98. doi: 10.1096/fj.11-201111. Epub 2012 Mar 29. FASEB J. 2012. PMID: 22459152 Free PMC article.
-
Tachykinins Processing is Significantly Impaired in PC1 and PC2 Mutant Mouse Spinal Cord S9 Fractions.Neurochem Res. 2015 Nov;40(11):2304-16. doi: 10.1007/s11064-015-1720-0. Epub 2015 Sep 15. Neurochem Res. 2015. PMID: 26373413
-
Role of pGlu-serpinin, a novel chromogranin A-derived peptide in inhibition of cell death.J Mol Neurosci. 2011 Oct;45(2):294-303. doi: 10.1007/s12031-011-9521-7. Epub 2011 May 3. J Mol Neurosci. 2011. PMID: 21537909 Free PMC article.
-
Peptidome analysis of umbilical cord mesenchymal stem cell (hUC-MSC) conditioned medium from preterm and term infants.Stem Cell Res Ther. 2020 Sep 23;11(1):414. doi: 10.1186/s13287-020-01931-0. Stem Cell Res Ther. 2020. PMID: 32967723 Free PMC article.
References
-
- Holz RW, Fisher SK. Synaptic transmission and cellular signaling: an overview. In: Siegel GJ, editor. Basic neurochemistry. Philadelphia: Lippincott Williams & Wilkins; 1999. pp. 191–212.
-
- Law PY, Loh HH. Regulation of opioid receptor activities. J Pharmacol Exp Ther. 1999;289:607–624. - PubMed
-
- Norris D. Vertebrate endocrinology. San Diego: Academic; 1997.
-
- Vaudry H. Brain peptides. In: Kastin AJ, editor. Handbook of biologically active peptides. Amsterdam: Elsevier; 2006. pp. 621–828.
-
- Krieger D, Brownstein MJ, Martin JB. Brain peptides. New York: Wiley-Interscience; 1983.
Publication types
MeSH terms
Substances
Grants and funding
- R01 DA004271/DA/NIDA NIH HHS/United States
- P41RR024851/RR/NCRR NIH HHS/United States
- R01NS24553/NS/NINDS NIH HHS/United States
- R01MH077305/MH/NIMH NIH HHS/United States
- R01 NS024553/NS/NINDS NIH HHS/United States
- P01HL58120/HL/NHLBI NIH HHS/United States
- R01DA04271/DA/NIDA NIH HHS/United States
- P41 RR024851/RR/NCRR NIH HHS/United States
- R01 RR016522/RR/NCRR NIH HHS/United States
- P01 HL58120/HL/NHLBI NIH HHS/United States
- R01 MH077305/MH/NIMH NIH HHS/United States
- P01 HL058120/HL/NHLBI NIH HHS/United States
- R01RR016522/RR/NCRR NIH HHS/United States
