

Moesin1 and Ve-cadherin are required in endothelial cells during in vivo tubulogenesis
- PMID: 20736288
- PMCID: PMC2926960
- DOI: 10.1242/dev.048785
Moesin1 and Ve-cadherin are required in endothelial cells during in vivo tubulogenesis
Abstract
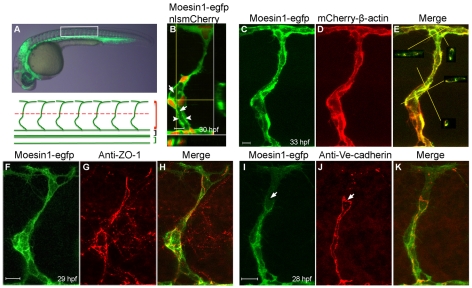
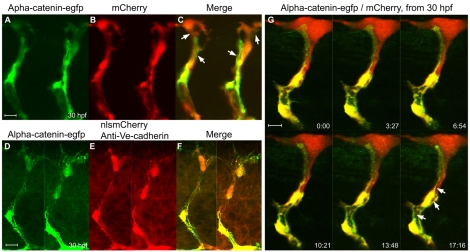
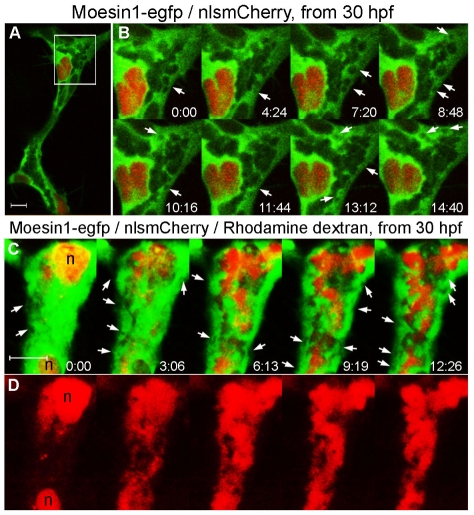
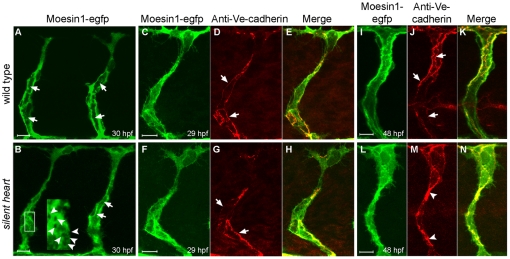
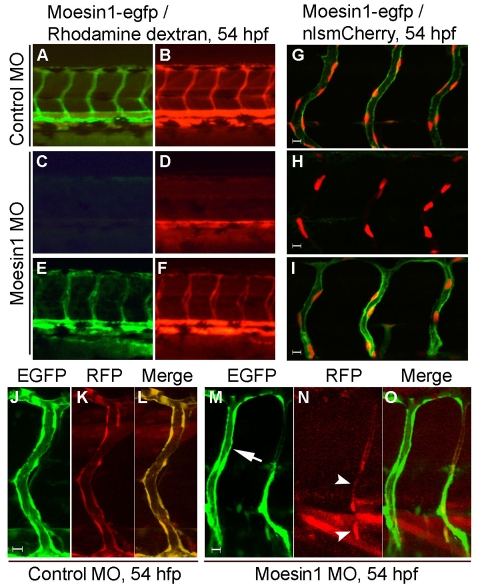
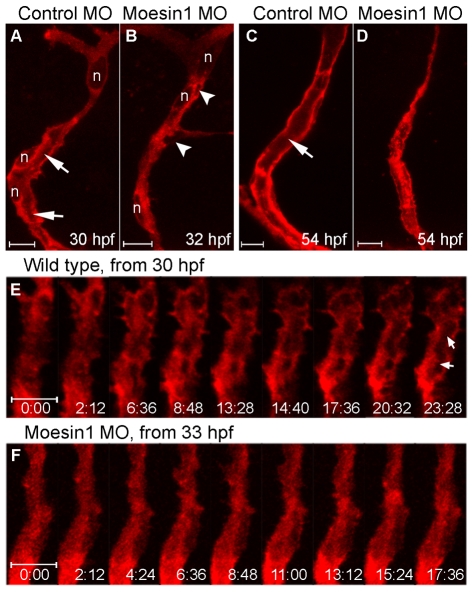
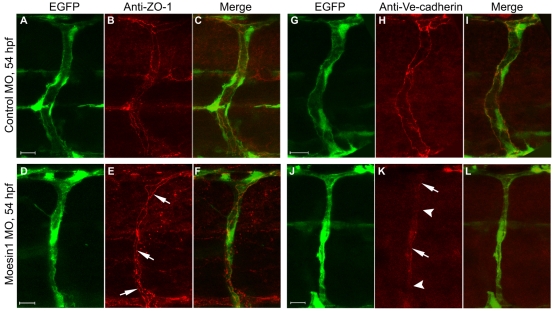
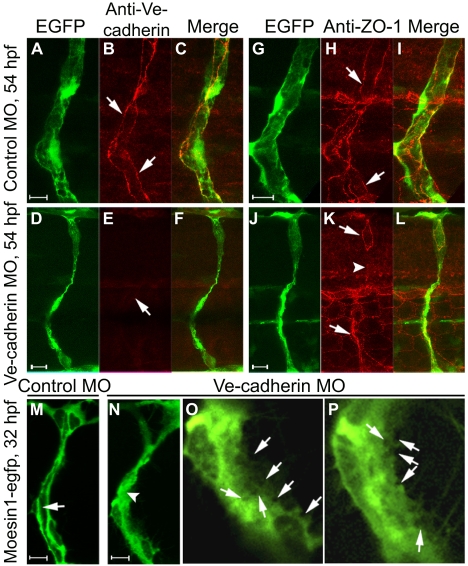
Endothelial tubulogenesis is a crucial step in the formation of functional blood vessels during angiogenesis and vasculogenesis. Here, we use in vivo imaging of living zebrafish embryos expressing fluorescent fusion proteins of beta-Actin, alpha-Catenin, and the ERM family member Moesin1 (Moesin a), to define a novel cord hollowing process that occurs during the initial stages of tubulogenesis in intersegmental vessels (ISVs) in the embryo. We show that the primary lumen elongates along cell junctions between at least two endothelial cells during embryonic angiogenesis. Moesin1-EGFP is enriched around structures that resemble intracellular vacuoles, which fuse with the luminal membrane during expansion of the primary lumen. Analysis of silent heart mutant embryos shows that initial lumen formation in the ISVs is not dependent on blood flow; however, stabilization of a newly formed lumen is dependent upon blood flow. Zebrafish moesin1 knockdown and cell transplantation experiments demonstrate that Moesin1 is required in the endothelial cells of the ISVs for in vivo lumen formation. Our analyses suggest that Moesin1 contributes to the maintenance of apical/basal cell polarity of the ISVs as defined by adherens junctions. Knockdown of the adherens junction protein Ve-cadherin disrupts formation of the apical membrane and lumen in a cell-autonomous manner. We suggest that Ve-cadherin and Moesin1 function to establish and maintain apical/basal polarity during multicellular lumen formation in the ISVs.
Figures
References
-
- Bagnat M., Cheung I. D., Mostov K. E., Stainier D. Y. (2007). Genetic control of single lumen formation in the zebrafish gut. Nat. Cell Biol. 9, 954-960 - PubMed
-
- Bayless K. J., Davis G. E. (2002). The Cdc42 and Rac1 GTPases are required for capillary lumen formation in three-dimensional extracellular matrices. J. Cell Sci. 115, 1123-1136 - PubMed
-
- Bayless K. J., Davis G. E. (2004). Microtubule depolymerization rapidly collapses capillary tube networks in vitro and angiogenic vessels in vivo through the small GTPase Rho. J. Biol. Chem. 279, 11686-11695 - PubMed
-
- Berryman M., Franck Z., Bretscher A. (1993). Ezrin is concentrated in the apical microvilli of a wide variety of epithelial cells whereas moesin is found primarily in endothelial cells. J. Cell Sci. 105, 1025-1043 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
